




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
干旱对桑树叶片光系统Ⅱ活性的影响
[滕志远1  , 张会慧
, 张会慧2 , 代欣1 , 胡举伟1 , 张秀丽1, * , 孙广玉1 ]
, 张会慧, 孙广玉|
|


作者简介:滕志远(1992—),男,山东招远人,硕士研究生,研究方向为植物生物科学。E-mail:1005066076@qq.com
栽植桑树既有防风固沙的生态价值,又具有增加农民收入的多种经济效益。随着国家“东桑西移,南蚕北移”工程的实施,栽桑业逐渐由湿润区域向干旱地区转移。干旱胁迫对于桑树光系统Ⅱ(PSⅡ)活性影响的生理机制尚不明确,该研究利用快速叶绿素荧光技术,研究干旱胁迫对桑树叶片PSⅡ活性的影响。结果表明:干旱胁迫导致叶片光系统Ⅱ活性受到明显的抑制,桑树叶片光反应中心潜在活性( Fv/ Fo)和以光吸收为基础的光合性能指数( PIABS)均显著低于对照,但干旱协迫对桑树叶片最大光化学效率( Fv/ Fm)的影响不显著。干旱处理的桑树叶片PSⅡ电子受体侧的电子由QA向QB传递受阻,同时PQ库和QB的电子接受能力下降,进而增加了
Planting mulberry ( Morus alba) not only had ecological value including windbreak and sand-fixation, but also had a variety of economic benefits, such as increasing farmers' income. With the implementing of “mulberry should be planting from southeast to northwest region”, the industry of mulberry is moving from the wet area to arid regions. The physiological mechanism of drought stress on photosystemⅡ (PSⅡ) photochemical activity have been not clear up to now, so effects of drought stress on PSⅡ photochemical activity in mulberry leaves were investigated with chlorophyll a fluorescence transient (OJIP curve) in this study. The results showed that potential photochemical efficiency of PSⅡ ( Fv/ Fo) and photosynthetic performance index ( PIABS) based on light absorption were significantly lower than the control, which indicated that photo-activity of leaf could be inhibited by drought stress. Maximal photochemical efficiency of PSⅡ ( Fv/ Fm) did not have a remarkable change. The electron transmission from QA to QB in PSⅡ electronic receptor side was relatively decreased in normalized OJIP curve, and electron-accepting ability of PQ electronic library and QB were also decreased, which increased the accumulation of
桑树是桑科(Moraceae)桑属(Morus L.)植物, 用途广泛。桑叶除了作为家蚕的主要饲料外, 还可以作为畜禽优质饲料源, 桑葚可开发为功能性食品和保健品, 而桑根、皮等含有多种活性物质, 可入药。桑树的抗逆性和适应性较强, 在盐碱地、荒山、沙漠及荒漠等地区均可种植, 具有重要的生态价值[1]。随着国家“ 东桑西移, 南蚕北移” 工程的实施, 栽桑业从内陆湿润区域向内陆干旱和半干旱地区转移[2]。干旱是内陆干旱地区常见的环境胁迫因子之一, 严重影响农林业植物的生长和发育, 限制农林业的发展。
植物的光合作用对干旱胁迫反应较为敏感, 可以通过检测干旱胁迫条件下植物的光合参数变化情况来鉴别植物的抗旱能力[3, 4], 植物净光合速率的大小取决于1, 5-二磷酸核酮糖羧化酶(Rubisco)的活性和电子传递速率[5]。桑树叶片的光合气体交换参数对土壤水分含量的变化具有明显的阈值响应, 有利于桑树光合作用和维持高水分利用效率的适宜土壤相对含水量为45%~78% [6]。而植物叶片光系统Ⅱ (PSⅡ )活性的降低被认为是植物发生光抑制的重要原因之一, PSⅡ 也是植物发生光抑制时的重要破坏位点, PSⅡ 作为植物类囊体膜上电子传递链的主要组成部分, 其活性与功能直接影响到NADPH和ATP的合成, 还会间接导致活性氧(ROS)的大量产生, 进一步导致光合能力的降低[7]。大量的研究表明, 在逆境条件下许多植物因其PSⅡ 功能降低或PSⅡ 结构损伤而使同化物积累减慢、产量降低[8, 9]。胡杨在干旱胁迫下, 当叶片的PSⅡ 尚未发生不可逆的光损伤时, 用于光合作用的光能所占的份额就明显下降, 耐受高光强的能力减弱, 捕获的光能过剩程度加剧, 即由此带来的光系统损伤的潜在危险增加, 进而发生光抑制[10]。闫海霞等[11]研究发现, 干旱胁迫使条墩桑叶片PSⅡ 潜在活性中心受损, 使其原初光能转换效率明显下降, 同时色素吸收的光能用于光合作用的比例显著下降, 而通过热和荧光形式散失的比例明显增加。有研究发现在干旱条件下, 印度桑树品种(Morus indica cv.V1)叶片PSⅡ 的结构和功能仍然保持完整, 而PSⅡ 电子传递受阻, 热耗散增强[12]。
目前, 有关干旱胁迫对于桑树叶片光合作用影响的生理机制鲜见报道。叶绿素荧光诱导动力学曲线(OJIP)可以快捷无损伤地分析逆境对光合作用某一过程的具体影响[13, 14]。为此, 本研究采用快速OJIP技术, 研究重度干旱对桑树叶片的PSⅡ 光化学活性、电子传递、放氧复合体(OEC)活性的影响机制, 其结果可为内陆干旱地区的桑树栽培提供参考。
供试桑树品种为一年生秋雨桑(Morus alba L.cv Qiuyu)的实生苗, 由黑龙江省蚕业研究所提供。试验于2014年5月在东北林业大学植物营养实验室进行。为确保试验材料的相对一致性, 在移栽之前将幼苗主根和主茎各保留5 cm, 然后将幼苗移栽到直径16 cm、高20 cm的培养钵中, 每钵定植3株, 总计90株。
为排除培养土差异影响和便于干旱处理, 将培养苗木的草炭土均匀混合后, 再与蛭石按1:1混合装盆, 待幼苗培养至30 d, 长出6~7片叶时, 选择长势一致的桑树幼苗进行试验。因为桑树极其耐旱, 预试验结果显示中度胁迫与对照没有差异, 因此本研究选择土壤相对含水量小于阈值的重度干旱(40%± 5%)作为处理。处理前使各盆的土壤相对含水量基本达到饱和, 停止浇水, 自然干旱, 通过称盆质量监测处理失水情况, 每天早晚用电子秤(精度5 g)称重, 并浇水至土壤相对含水量分别达到80%(对照组, CK)和40%± 5%(处理组, 重度干旱)[15]。持续干旱胁迫10 d后, 对桑树幼苗进行快速叶绿素荧光动力学曲线的测定。
OJIP的检测:于2014年6月18日上午9:00~11:00, 此时段天气晴朗, 分别选择对照组和处理组中长势相对一致的5株幼苗(5次重复), 将桑树幼苗从上往下数第4片完全展开叶进行40 min的暗适应后, 利用Mini调制式掌上叶绿素荧光仪(FluorPen FP 100 max, 捷克)测定叶片的OJIP。在测定中, 诱导光强设为3 000 μ mol· m-2· s-1的脉冲红光, 从10 μ s开始记录相对荧光强度(Fv)共记录1 s, OJIP曲线上O, J, I和P点对应的时刻分别为20, 2 000, 30 000和1 000 000 μ s, 利用5次重复的平均值绘制OJIP曲线。
O, L, K, J, I和P点的相对荧光强度分别以Fo, FL, FK, FJ, FI和FP表示, 而L, K, J和I点的相对可变荧光分别以VL, VK, VJ和VI表示, 其中K点和L点分别为OJIP曲线上0.3 ms和0.15 ms对应的时刻。OJIP曲线是按照Strasser等[16]的方法利用JIP-test进行分析, 特异分析J点和I点可变荧光的变化, 将O-P曲线标准化, 即将O点的相对荧光强度(Fo)设为0, 而P点相对荧光强度(Fp)设为1, 按公式(Ft-Fo)/(Fm-Fo)求得各时间点的相对可变荧光, 其中Ft表示各时间点的相对荧光强度。为研究干旱胁迫对桑树叶片放氧复合体OEC活性的影响, 即K点和L点的相对可变荧光VL和VK变化, 分别将O-J和O-K曲线进行标准化, 其计算公式分别为:(Ft-Fo)/(FJ-Fo)和(Ft-Fo)/(FK-Fo), 计算干旱胁迫处理组桑叶标准化O-J和O-K曲线与对照组之间的差值, 分别以△ VO-J, △ VO-K表示。
叶片相对含水量的测定:采新鲜叶片测定鲜重后, 将叶片浸入蒸馏水饱和吸水24 h, 称取饱和重, 然后烘干测定干重, 相对含水量=(鲜重-干重)/(饱和重-干重)[17]。叶绿素含量的测定参照王晶英[18]的方法。
运用DPS软件对试验数据进行统计分析, 利用Excel软件绘制图表, 图中数据为5次重复的平均值± 标准差(SD), 采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)进行差异显著性分析。
供试桑树叶片情况见图1, 对照组桑树叶片平展上举、颜色鲜绿(图1-A), 干旱处理组桑树幼苗叶片萎蔫下垂、颜色浅绿(图1-B)。
 | 图1 测定前桑树幼苗叶片状态Fig.1 Growth condition of mulberry leaves before measuring |
由图2可以看出, 处理组叶片的相对含水量在70%左右, 极显著低于对照(80%左右)(P< 0.01)。干旱处理10 d后, 与对照组相比, 处理组叶片的总叶绿素Chl (a+b)、叶绿素a(Chl a)和叶绿素b(Chl b)含量均明显降低, 分别降低了35.93%, 30.87%和45.12%(P< 0.01), 其中Chl b下降的程度较Chl a大, 干旱处理的Chl a/Chl b较对照组高25.27%(P< 0.01)。
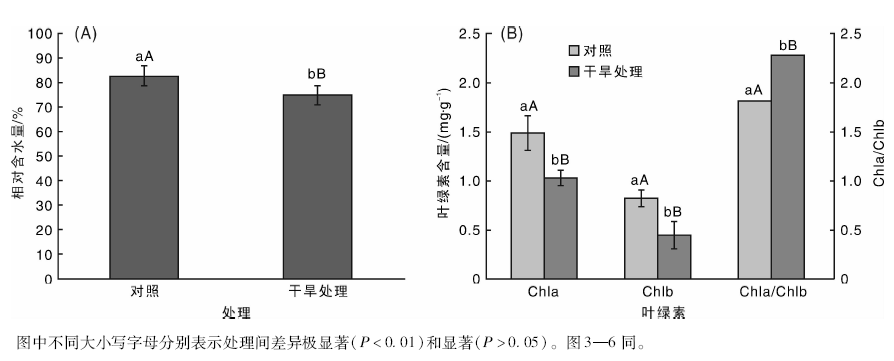 | 图2 干旱对桑树叶片相对含水量和叶绿素含量的影响Fig.2 Effects of drought stress on relative water content and chlorophyll content in mulberry leaves |
由图3可以看出, 在干旱胁迫下, 代表反应中心PSⅡ 潜在活性的荧光参数Fv/Fo比对照组低7.9%, 差异极显著(P< 0.01); 最大光化学效率Fv/Fm较对照组略低, 但差异不显著(P> 0.05), 而以光吸收为基础的光合性能指数(PIABS)较对照组低35%, 差异极显著(P< 0.01)。
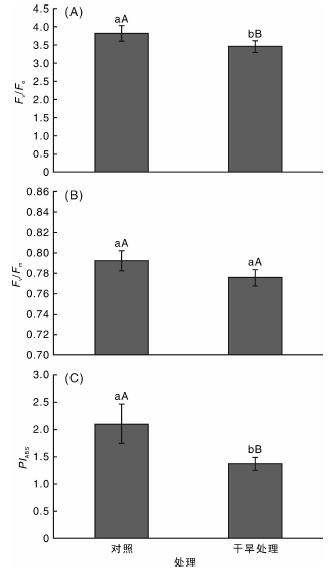 | 图3 干旱对桑树叶片光化学效率Fv/Fo, Fv/Fm和PIABS的影响Fig.3 Effects of drought stress on Fv/Fo, Fv/Fm and PIABS in mulberry leaves |
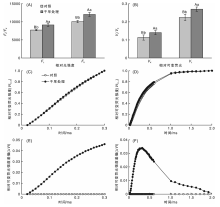
由图4可以看出, 干旱胁迫明显改变了桑树叶片的OJIP曲线形态, 其中O点、J点、I点和P点的相对荧光强度(Fo, FJ, FI和FP)明显升高。定量分析干旱胁迫对OJIP曲线各点相对荧光强度的影响, 结果表明, 干旱胁迫下桑树叶片的Fo较对照高13.94%, 差异极显著(P< 0.01), 而FP较对照上升了6.61%, 差异不显著(P> 0.05)。同时, 干旱处理使桑树叶片的FJ和FI分别较对照升高了16.93%和8.64%, 处理之间差异极显著(P< 0.01)。
 | 图4 干旱胁迫对桑树叶片OJIP曲线的影响Fig.4 Effects of drought stress on OJIP transient in mulberry leaves |
由图5可以看出, 干旱胁迫增加了OJIP曲线上L点和K点的相对荧光强度(FL和FK), FL和FK分别较对照组升高18.39%(P< 0.01)和19.64%(P< 0.01), 差异极显著。分别将O-K和O-J曲线标准化后可以发现, 干旱胁迫处理的桑树叶片标准化O-K曲线上0.15 ms时即L点的相对可变荧光(VL)和标准化O-J曲线上0.3 ms时即K点的相对可变荧光(VK)均与对照组之间有明显差异, 分别将标准化O-K和O-J曲线与对照组曲线作差值分析, 可以看出L点和K点的相对可变荧光与对照组之间差异最大。定量分析结果表明, 干旱胁迫下桑树叶片的VL和VK分别较对照组升高了16.07%和23.62%, 差异均极显著。
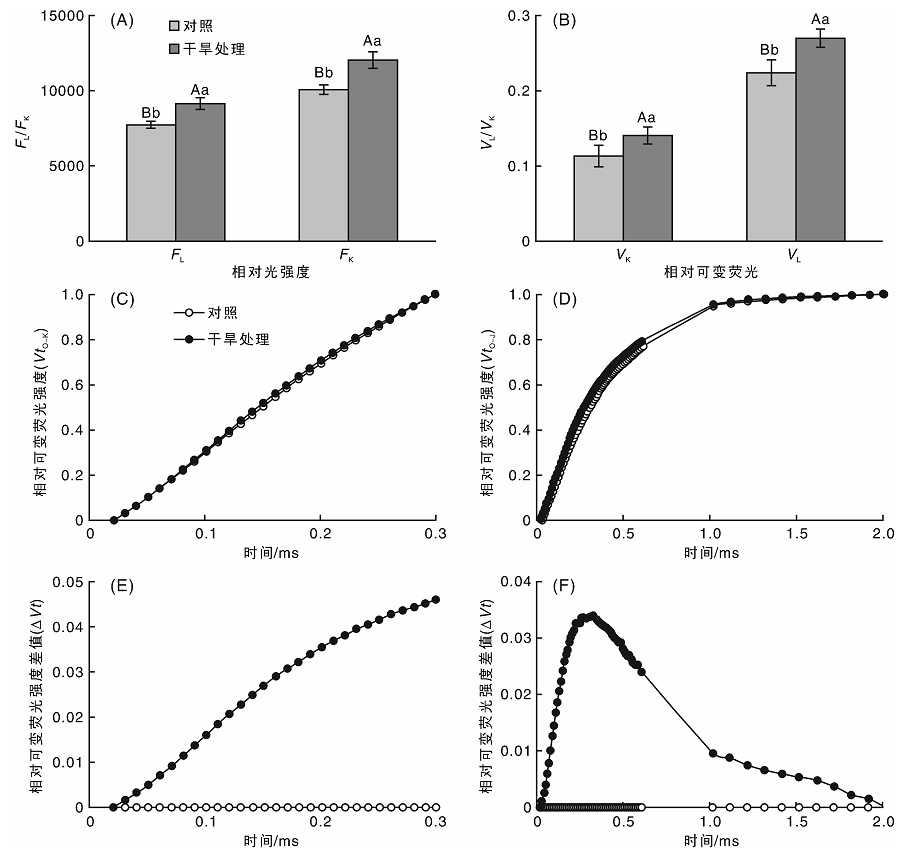 | 图5 干旱胁迫对桑树叶片标准化OJ和OK曲线的影响Fig.5 Effects of drought stress on normalized OJ and OK transient in leaves of mulberry |
图6可以看出, 将OJIP曲线按O-P标准化后可以发现, 干旱胁迫增加了桑树叶片J点和I点的相对可变荧光(VJ和VI)。将干旱胁迫桑树叶片标准化O-P曲线与对照做差值可以发现, 干旱胁迫使VJ的增加幅度大于VI。定量分析结果表明, 干旱胁迫下桑树叶片VJ和VI分别增加了34.15%(P< 0.01)和20.47%(P< 0.01)。
 | 图6 干旱对桑树叶片标准化OJIP曲线的影响Fig.6 The effects of drought on normalized OJIP transient in leaves of mulberry |
光合作用是植物获得物质和能量的重要代谢过程之一[19, 20], 其经常受到外界条件变化的影响。光合作用可作为植物长势和抵抗逆境能力的参考指标[21], 植物水分供应情况直接影响其光合作用能力, 水分亏缺导致光合能力下降, 是干旱条件下农林业植物减产的主要原因之一[22]。叶绿素荧光被称为光合作用的探针, 叶绿素荧光诱导动力学曲线可以反映环境因子对光合作用过程的影响[23], 目前已证明活体叶绿素荧光主要与PSⅡ 有关。
本研究表明, 在导致桑树叶片持续萎蔫的重度干旱胁迫下, 桑树叶片中Chl (a+b), Chl a和Chl b含量以及PIABS, Fv/Fo均明显低于对照组(P< 0.01), 而Fv/Fm虽然较对照组低, 但差异不显著(P> 0.05)。Fv/Fm通常被作为逆境下PSⅡ 光化学反应状况的叶绿素荧光参数, 大多数植物叶片在非光抑制条件下的Fv/Fm约在0.8左右, 物种之间差异较小。Christen等[24]、Oukarroum等[25]和Guha等[12]发现, 桑树的Fv/Fm对干旱胁迫不敏感, 原因在于该参数值的大小取决于Fo和Fm, 如果这两者发生相同趋势的变化, 其比值变化就不明显。在本试验中, 干旱胁迫下Fo和Fm均表现出上升的趋势, 所以Fv/Fm变化不明显。
PIABS不但可以反映原初光化学量子产率, 而且反映了反应中心密度和电子在PSⅠ 和PSⅡ 之间的传递情况, 可以从光能吸收、捕获和电子传递3方面综合反映光系统的活性, 敏感性大于Fv/Fm[26-27]。为此, 可将PIABS作为本试验供试桑树品种对干旱胁迫反应的有效荧光参数指标。Fo是PSⅡ 反应中心完全开放时的荧光产量, 其值的大小取决于PSⅡ 天线色素内的最初激发态电子密度、激发能传递速率和光合色素浓度[28]; 而FP是PSⅡ 反应中心完全关闭时的荧光产量, 此时反应中心不再接受光量子, 其大小可反映PSⅡ 中电子传递情况[29]。綦伟等[30]研究发现, 在干旱胁迫下, 葡萄砧木Fo上升而FP下降, 其中抗旱性强的品种Fo和FP变化的幅度较小, PSⅡ 光化学活性强。在本试验中, 干旱胁迫的桑树Fo较对照高, 而FP与对照差异不显著, 说明干旱胁迫导致桑树叶片PSⅡ 活性降低。
为分析干旱胁迫对桑树光合作用的具体影响, 本试验将桑树叶片的OJIP曲线按O-P标准化后特异分析了L, K, J和I点的相对可变荧光变化。OJIP曲线上K点(0.3 ms)相对可变荧光VK的增加是放氧复合体OEC受到伤害的特异性标志[31, 32], 而OJIP曲线上L点(0.15 ms)相对可变荧光VL的上升则是类囊体解离的特异性标志[33]。本试验分别将O-J和O-K曲线标准化后分析其K点和L点相对可变荧光的变化, 发现在干旱胁迫下, 桑树叶片的VK和VL均明显升高, 说明干旱胁迫抑制了放氧复合体OEC的活性, 可能导致类囊体发生解离, 减弱了PSⅡ 蛋白复合体亚单位之间的连接紧密性, 抑制PSⅡ 的光化学活性, 增加光抑制程度。OJIP曲线上J点(2 ms)的相对可变荧光VJ表示关闭反应中心的数量或QA的还原量, 即VJ呈增加趋势说明
综上所述, 水分极度亏缺时, 桑树幼苗叶片的PSⅡ 反应中心活性显著下降, 光抑制程度明显。其具体的阻遏原因在于干旱减弱了PSⅡ 受体侧的电子传递能力, 特别是抑制了QB接受电子的能力; 降低了PSⅡ 供体侧放氧复合体OEC的活性, 加快了类囊体的解离速度, 进而破坏PSⅡ 的正常生理功能。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|