
{kind=link}
{kind=link}
{kind=link}
C3和C4植物光能利用效率和水分利用效率的比较研究
[叶子飘1  , 杨小龙
, 杨小龙1 , 康华靖2 ]
, 杨小龙|
|


作者简介:叶子飘(1964—),男,江西永新人,博士、教授,主要从事植物光合作用对光响应模型研究。E-mail:yezp@igsu.edu.cn
了解C3和C4植物光能利用效率(LUE)和水分利用效率(WUE)对光的响应关系,对揭示植物的光能利用与需水规律具有重要意义。以栾树( Koelreuteria paniculata)、辣椒( Capsicum annuum)、高粱( Sorghum bicolor)和玉米( Zea mays)为研究对象,用LI-6400光合仪测量了这4种植物的气体交换参数,并利用光响应机理模型所构建的叶片光能和水分利用效率对光的响应模型分别研究了它们的光能和水分利用效率的变化规律。结果表明:(1)栾树和辣椒的最大光能利用效率(LUEmax)和最大水分利用效率(WUEmax)比高粱和玉米低,所对应的饱和光强( IL-sat)也小于高粱和玉米;(2)高粱的最大净光合速率( Pnmax)和饱和光强( Isat)均比玉米小,但它的LUEmax和WUEmax却比玉米大。由此可知,C3植物的栾树和辣椒LUEmax和WUEmax比C4植物的高粱和玉米低,而相应的饱和光强( IL-sat和 IW-sat)则没有这种趋势。
This study aims to investigate the response relationship between light-use efficiency (LUE), water-use efficiency (WUE) and light intensity for C3 and C4 plants species. It is an important significance to reveal light-use and water consume of plants. We used LI-6400 Portable Photosynthetic System to measure the leaf gas exchange data of Koelreuteria paniculata, Capsicum annuum, Sorghum bicolor and Zea mays. Then these data were simulated by a light-response model of LUE and WUE which were developed by a mechanistic model of light-response of photosynthesis. These results showed that: (1) the maximum light-use efficiency (LUEmax), maximum water-use efficiency (WUEmax) and the corresponding saturation irradiance ( IL-sat for LUEmax) of K. paniculata and C. annuum were less than that of Z. mays and S. bicolor; (2) the maximum net photosynthetic rate ( Pnmax) and saturation irradiance ( Isat for Pnmax) of S. bicolor were less than that of Z. mays, but its LUEmax and WUEmax were more than that of Z. mays. Therefore, LUEmax and WUEmax of K. paniculata and C. annuum for C3 plants are less than that of S. bicolor and Z. mays for C4 plants. However, there is no such tendency in their corresponding saturation irradiances (i.e. IL-sat and IW-sat) for four plants.
光能利用效率(light-use efficiency, LUE)是植物光合作用量子效率的固定参数, 被定义为叶片的净光合速率(Pn)与其所吸收光合光量子通量密度(photosynthetic photon flux density, PPFD)的比值[1, 2, 3], 其数学表达式为LUE=Pn/PPFD, 单位为mol· mol-1或μ mol· μ mol-1。如今, 光能利用效率这个参数被广泛用于评价植物叶片甚至整个冠层对光能的利用能力上[2, 3, 4, 5, 6, 7, 8, 9, 10]。与光能利用效率相对应的另一个重要参数是水分利用效率(water-use efficiency, WUE), 它是耦合植物叶片光合与水分生理过程的重要指标, 是表征植物在等量水分消耗情况下, 固定CO2的能力, 是植物叶片水分利用特征的基本生理参数[11], 其数学表达式为WUE=Pn/Tr, 单位为μ mol· mmol-1。植物叶片水平的WUE研究不仅可揭示植物叶片内在的耗水机制, 明确植物自身光合能力大小, 还能反映植物有效利用水分的能力, 为人们了解植物水分利用特征提供有用的信息, 因而具有重要的指导应用价值。
人们进行了大量的C3和C4植物光合作用和蒸腾作用对不同生境响应的对比实验研究[12, 13, 14, 15, 16, 17, 18], 试图揭示这些植物的光合特性、光能利用效率和水分利用效率对物种分布的影响。绝大多数关于C3和C4植物生理生态的研究表明, 两者的光合特性存在较大的差异[12, 13, 14, 15, 16]。相对于C3植物, C4植物的光能利用效率高, 这是因为C4植物光合作用是由维管束鞘细胞和叶肉细胞两种细胞联合起来共同完成的, C4植物CO2同化率高, 而气孔对水蒸气导度较小, 蒸腾失水较少, 因而具有更高的水分利用效率[12, 15, 18]和光能利用效率[14, 15, 16, 18]。关于C3植物和C4植物叶片LUE和WUE对光强的响应规律尚不明晰, 也不清楚在一定环境条件下照射到植物叶片的光合光量子通量密度多大时它们的LUE和WUE能达到最大值。此外, 对C3和C4植物光能利用和水分效率的变化规律是否相同, 是否存在差异等问题也不了解。而这些问题的探索对正确评价不同环境下植物叶片的光能和水分利用效率是非常重要的。
本研究将首先以Ye等[19]的机理模型为基础构建叶片的光能利用效率和水分利用效率与光强之间的关系式(简称为光能利用效率模型:LUE-I模型; 水分利用效率模型:WUE-I模型); 其次, 分别利用叶片的LUE-I和WUE-I模型得到植物叶片的最大光能利用效率(LUEmax)和水分利用效率(WUEmax)及与之对应的饱和光强(IL-sat是与LUEmax相对应, IW-sat是与WUEmax相对应); 最后, 以栾树(Koelreuteria paniculata)、辣椒(Capsicum annuum)、高粱(Sorghum bicolor)和玉米(Zea mays)为研究对象, 利用所构建叶片的LUE-I和WUE-I模型研究这4种植物叶片的光能利用效率和水分利用效率的变化规律, 以期为比较研究C3和C4植物的光能利用效率和水分利用效率等提供数学工具和理论依据。
植物叶片的光能利用效率是由植物光合作用对光响应的机理模型[19]得到的。而光响应机理模型的简化模型就是作者在2007年[20]构建的光响应新模型, 即:
Pn=α
式(1)中, α 是光响应曲线的初始斜率, β 和γ 分别为抑制系数和饱和系数, α , β 和γ 三个系数的具体表达形式见文献[19], I为光合光量子通量密度, Rd为暗呼吸速率。而植物叶片的饱和光强(Isat)和最大净光合速率(Pnmax)可以分别由Isat=
由LUE的定义, 利用式(1)就可以得到植物叶片的LUE为:
LUE=
如果用IL-sat表示与叶片最大光能利用效率(LUEmax)相对应的饱和光强, 那么它可以由式(3)得到, 即:
IL-sat=
把式(3)代入式(2)就可以得到
LUEmax=α
此外, 利用式(1)还可以得到植物叶片的水分利用效率为:
WUE=
式(5)中的KW=Rd/Tr。如果用IW-sat表示与叶片最大水分利用效率相对应的饱和光强, 那么它可以由式(6)得到, 即:
如果用WUEmax表示叶片的最大水分利用效率, 则把式(6)代入式(5)就可以得到WUEmax为:
WUEmax=α 1
栾树(Koelreuteria paniculata Laxm.)于2010年3月从温州泰顺白云园艺种苗场移植到温州职业科技学院校园的实验地; 辣椒(Capsicum annuum L.) 品种都椒1号种子购于温州田园种子公司, 于2010年3月17日播种, 4月下旬移栽; 玉米和高粱种子则于2010年3月直接撒播于温州职业科技学院校园的实验地。进行常规管理。于2010年6月份的晴天(此时植株处于旺盛生长期), 随机选择5株长势较为一致的健壮植株, 对4种植物叶片的光响应曲线进行测量, 测量结果取平均值, 详细的管理和实验见文献[19]。
自然光诱导1.0~1.5 h后, 采用开放式气路, 设定温度为30 ℃, 空气相对湿度为50%~70%, 应用LI-6400-40B提供不同的光合光量子通量密度(PPFD, μ mol· m-2· s-1), 分别在PPFD为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、150、100、50和0 μ mol· m-2· s-1和CO2浓度为380 μ mol· mol-1时测定这4种植物的光响应曲线。然后用光响应机理模型[19]拟合光响应曲线, 由此得到植物叶片的光合参数, 如Pnmax(最大净光合速率)、α (初始量子效率)、Rd (暗呼吸速率)、Ic (光补偿点) 、Isat (饱和光强)、β (抑制系数)和γ (饱和系数)。然后用式(3)和式(4)分别得到IL-sat和LUEmax; 用式(6)和式(7)分别得到IW-sat和WUEmax。
光合作用对光响应曲线的实验数据用由叶子飘提供的光合计算软件(光合计算软件4.1.1版本)处理, 叶片的光能利用效率对光响应曲线的实验数据处理用SPSS12.5版本(SPSS, Chicago, IL)处理; 作图由Origin7.0完成, 然后用Adobe Illustrator CS5进行图形处理, 采用SPSS 12.5做方差差异显著性分析。
图1为栾树、辣椒、高粱和玉米的光响应曲线。由图1-a可知, 栾树在750 μ mol· m-2· s-1左右就到达一个平台区, 然后随光强的增加净光合速率几乎不增加; 而辣椒则随光强的增加, 其净光合速率持续增加, 在所设定的测量光强范围之内尚没有光饱和现象出现(图1-b); 由图1-c可知, 高粱在高光强时其净光合速率随光强的增加而下降, 出现光抑制现象; 而玉米在高光强时其净光合速率略有下降(图1-d)。此外, 在光强小于1 000 μ mol· m-2· s-1时高粱的净光合速率与玉米几乎遵循相同的响应规律, 但在光强超过1 000 μ mol· m-2· s-1时, 其净光合速率随光强的增加而下降(图1-c), 而玉米则随光强的增加而增加(图1-d)。
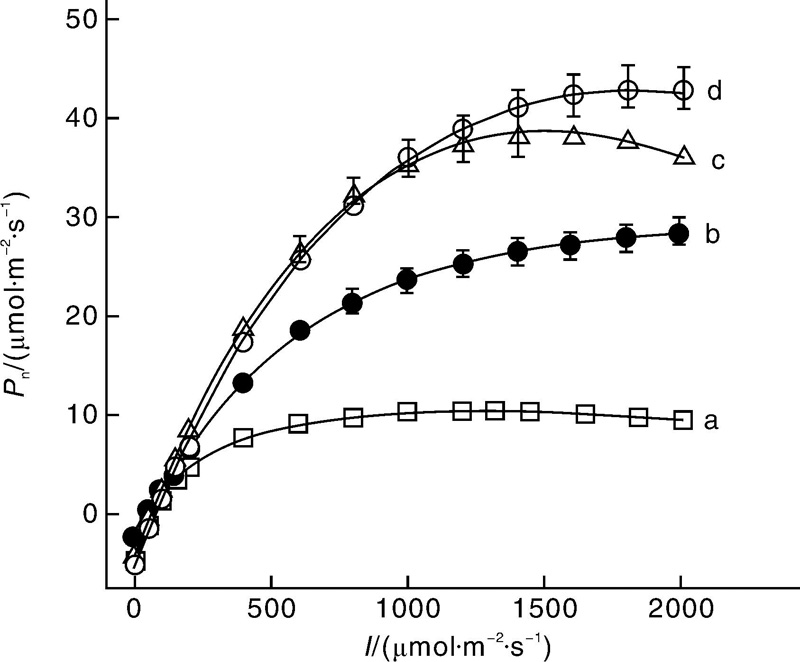 | 图1 栾树(a)、辣椒(b)、高粱(c)和玉米(d)的光响应曲线[19]Fig.1 Pn-I curves for Koelreuteria paniculata(a), Capsicum annuum(b), Sorghum bicolor(c) and Zea mays(d) |
表1给出了用公式(1)拟合栾树、辣椒、高粱和玉米的光响应曲线所得到的主要光合参数。由表1可知, C4植物的最大净光合速率远大于C3植物, 最小是栾树为10.36 μ mol· m-2· s-1, 最大是玉米为42.81 μ mol· m-2· s-1。但在本试验中, 高粱和玉米的饱和光强却比辣椒低, 且高粱出现明显的光抑制现象。
| 表1 栾树、辣椒、高粱和玉米的光合参数[19] Table 1 Photosynthetic parameters of Koelreuteria paniculata, Capsicum annuum, Sorghum bicolor and Zea mays |
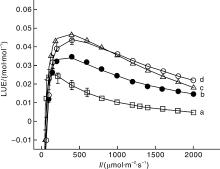
图2是栾树、辣椒、高粱和玉米的光能利用效率的光响应曲线。由图2-a和2-b可知, C3植物栾树和辣椒的光能利用效率的光响应曲线相似。在低光强处, 其光能利用效率随光强的增加而增加, 并且很快就达到了最大值, 随后随光强的增加呈非线性下降; 由图2-c和2-d可知, C4植物高粱和玉米在低光强时光能利用效率的行为与栾树和辣椒的相似, 但在高光强处其光能利用效率随光强的增加呈近似直线下降, 且在较低光强时高粱的光能利用效率比玉米大, 但在高光强时(大约光强在1 000 μ mol· m-2· s-1)其光能利用效率却比玉米的小。
表2给出了4种植物的最大光能利用效率和对应的饱和光强。由表2可知, 栾树的最大光能利用效率最小, 为0.025 6 mol· mol-1, 高粱的最大光能利用效率最大, 为0.048 6 mol· mol-1。此外, 辣椒的光能利用效率在较高光强时明显大于栾树; 高粱和玉米的光能利用效率则差异较小。与最大光能利用效率相对应的饱和光强最高的是玉米为391.82 μ mol· m-2· s-1, 其次是高粱为338.02 μ mol· m-2· s-1, 最小的是栾树为148.25 μ mol· m-2· s-1。
 | 图2 栾树(a)、辣椒(b)、高粱(c)和玉米(d)光能利用效率的光响应曲线Fig.2 Light-response curves of light-use efficiency for Koelreuteria paniculata(a), Capsicum annuum(b), Sorghum bicolor(c) and Zea mays(d) |
| 表2 栾树、辣椒、高粱和玉米的最大光能利用效率和饱和光强 Table 2 The maximum light use efficiency and saturation irradiance for Koelreuteria paniculata, Capsicum annuum, Sorghum bicolor and Zea mays |
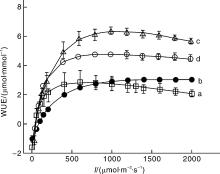
图3为4种植物的水分利用效率的光响应曲线。对比图3-a和3-b可知, 光强低于800 μ mol· m-2· s-1时栾树的水分利用效率比辣椒高, 光强高于800 μ mol· m-2· s-1时栾树的水分利用效率却比辣椒低, 且2种植物的水分利用效率差异不大; 对比图3-c和3-d可知, 在低光强(低于250 μ mol· m-2· s-1)时, 高粱和玉米的水分利用效率几乎相同, 但在高光强处高粱的水分利用效率明显高于玉米。
表3给出了4种植物的最大水分利用效率和对应的饱和光强。由表3的数据可知, 高粱的最大水分利用效率为6.36 μ mol· mmol-1, 其次是玉米为4.79 μ mol· mmol-1, 最小的是栾树为3.07μ mol· mmol-1。与最大水分利用效率对应的饱和光强, 最大的是辣椒为1 631.96 μ mol· m-2· s-1, 其次是高粱为1 109.25 μ mol· m-2· s-1, 最小是栾树为671.80 μ mol· m-2· s-1。
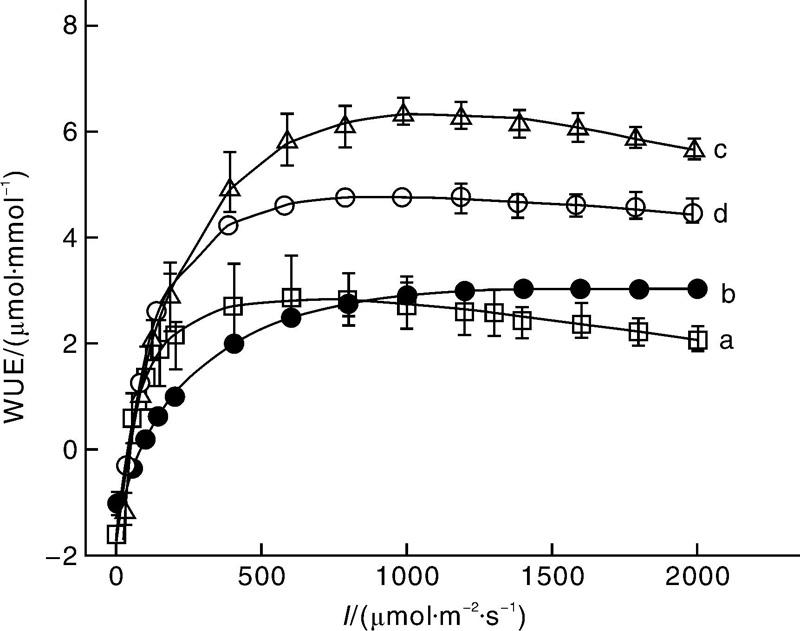 | 图3 栾树(a)、辣椒(b)、高粱(c)和玉米(d)水分利用效率的光响应曲线Fig.3 Light-response curves of water-use efficiency for Koelreuteria paniculata(a), Capsicum annuum(b), Sorghum bicolor(c) and Zea mays(d) |
| 表3 栾树、辣椒、高粱和玉米的最大水分利用效率和饱和光强 Table 3 The maximum water use efficiency and saturation irradiance for Koelreuteria paniculata, Capsicum annuum, Sorghum bicolor and Zea mays |
光能利用效率被用于评价植物叶片甚至整个冠层对光能的利用能力[2, 3, 4, 5, 6, 7, 8], 其值越大, 表明其利用光能的能力就越强。由表2的数据可知, 高粱和玉米的最大光能利用效率明显比栾树和辣椒的大, 表明高粱和玉米的光能利用效率较高, 同化CO2能力就更强, 即其净光合速率更大(表1)。由图2可知, 4种植物叶片的LUE在低光强时随光强的增加而快速增加至LUEmax; 当超过饱和光强之后光能利用效率的值随光强的增加而下降。但栾树和辣椒的光能利用效率的光响应曲线与高粱和玉米的略有差异, 在饱和光强之后高粱与玉米的光能利用效率随光强的增加几乎直线下降(图2), 而栾树和辣椒的光能利用效率在饱和光强之后随光强的增加而非线性下降, 与已有的研究结果类似[4, 5, 6, 7, 8, 9, 10], 不同之处表现在与LUEmax和相对应的饱和光强数值上的差异以及光能利用效率下降速率的不同。这种现象在其他植物中也存在, 例如, 于显枫等[21]发现, 高大气 CO2浓度(760 μ mol· mol-1)遮阴处理小麦叶片的最大LUE(LUEmax)大于正常大气CO2浓度(400 μ mol· mol-1)生长的小麦叶片, 且与LUEmax对应的饱和光强(IL-sat)也大于正常大气 CO2浓度生长的小麦; 王凯等[8]发现自然光和遮阴环境下黄波罗幼苗叶片在春节、夏季和秋季时IL-sat约在100 μ mol· m-2· s-1时叶片的LUE就达到最大值。孙景宽等[9]发现不同浓度盐分胁迫下二色补血草的光能利用效率达到最大值的IL-sat不同(约为200 μ mol· m-2· s-1)。在本研究中, 对比表1和表2的数据可知, 辣椒的净光合速率在光强在2 000 μ mol· m-2· s-1时尚没有达到最大值, 但它的LUE却在299.71 μ mol· m-2· s-1处就达到了最大值。玉米的最大净光合速率(Pnmax)和饱和光强(Isat)比高粱的值都大, 然而在光强小于1 000 μ mol· m-2· s-1时其LUE却比玉米大。由图1-c和1-d与图2-c和2-d可知, 光强为1 000 μ mol· m-2· s-1时是高粱和玉米两种植物利用光能能力出现差异的一个节点。
WUE是表征植物叶片水分利用特征的基本生理参数[12]。由图3可知, WUE的变化规律与LUE相似, 在较低的光强时植物叶片的WUE随光强的增加而非线性增加达到最大值, 然后它的值随光强的增加而下降, 这与已有的研究结果类似
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|