



{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同日龄仔猪外周血T淋巴细胞的检测与分析
[邹璐1, 2  , 张景艳
, 张景艳2 , 张凯2 , 王磊2 , 张康2 , 王旭荣2 , 王学智2 , 余四九1 , 杨志强2, * , 李建喜2, * ]
, 张景艳, 李建喜]
|
|


作者简介:邹璐(1990—),男,河南潢川人,硕士研究生,主要从事中兽药的研究。E-mail: 1406832500@qq.com
为建立健康仔猪外周血淋巴细胞亚群的临床参考范围,试验分析不同生长阶段仔猪的免疫系统功能水平,分别在健康仔猪断奶后第1、7、21和35天采取外周血,分离外周血单个核细胞,采用三色流式细胞术的方法检测各类主要T淋巴细胞亚群所占外周血单个核细胞的比例,及其占CD3+ T淋巴细胞亚群中的比例,计算CD4+ /CD8+ 的比值。结果表明,随着日龄的增加公母仔猪CD3+ T淋巴细胞亚群占外周血单个核细胞的比例逐渐升高,分别从(38.00±10.94)% 和(38.17±15.39)%上升至(55.43±12.30)%和(51.57±10.60)%。在断奶后1 d和7 d公母仔猪CD3+ T淋巴细胞比例无显著性差异( P>0.05),至断奶后21 d母仔猪CD3+ T淋巴细胞比例显著增加( P<0.05),公仔猪至35 d时CD3+ T淋巴细胞比例显著增加( P<0.05)。随着日龄的增长,公母仔猪CD4+ T淋巴细胞亚群占外周血单个核细胞的比例分别由(10.29±2.71)% 和(10.3±2.17)%上升至(22.2±5.87)%和(23.4±6.84)%。公母仔猪CD4+ T淋巴细胞的比例逐渐升高,且各个时间点公母仔猪结果无明显差异( P>0.05)。公母仔猪CD8+ T淋巴细胞占外周血单个核细胞的比例在各时间点分别为(23.80±6.67)%、(17.14±10.64)%、(25.46±12.32)%、(33.33±8.93)%和(28.55±6.75)%、(23.62±11.51)%、(37.37±13.18)%、(38.31±15.66)%。随着日龄的增加公母断奶仔猪CD8+ T淋巴细胞呈现先下降后升高的趋势。断奶仔猪各淋巴细胞亚群分布会随着断奶时间的增加而呈现不同的变化,并且存在着性别上的差异。结合两种不同的细胞圈定方法,能够更准确地反映出仔猪体内各淋巴细胞亚群数量和比例的变化。
, ZHANG Jingyan, LI Jianxi
To establish healthy piglets of peripheral blood lymphocyte subsets in clinical reference range, and analyze the level of immune system in different growth stages of piglets, the blood samples were collected on the 1st day, 7th day, 21st day and 35th day after weaning, respectively, and the mononuclear cells were isolated from the peripheral blood. Three color flow cytometry was used to detect the proportion of primary kinds of T lymphocyte subsets in peripheral blood mononuclear cells and in CD3+ T lymphocyte subsets, then the ratio of CD4+ /CD8+ was calculated. The proportion of CD3+ T lymphocyte subsets for peripheral blood mononuclear cells in male and female piglets were gradually increased from (38.00±10.94)% to (55.43±12.30)% and from (38.17±15.39)% to (51.57±10.60)% , respectively with the increase of age. On the 1st day and 7th day after weaning, the CD3+ T lymphocyte ratio had no significant difference between male and female piglets ( P>0.05). On the 21st day after weaning, the CD3+ T lymphocyte ratio showed significant increase in male piglets( P<0.05). On the 35th day, the percentage of CD3+ T cells showed a significant increase ( P<0.05). With the increase of age, the proportion of CD4+ T lymphocyte subsets in peripheral blood mononuclear cells of male and female piglets increased from (10.29±2.71)% to( 22.2±5.87)% and from (10.3±2.17)% to (23.4±6.84)%, respectively. The proportions of CD4+ T lymphocytes in male and female piglets were gradually increasing, and there was no significant difference between male and female piglets at different time points( P>0.05). The proportions of CD8+ T lymphocytes in peripheral blood mononuclear cells at the four time points were(23.80±6.67)%, (17.14±10.64)%, (25.46±12.32)%, (33.33±8.93)% in male piglets and (28.55±6.75)%, (23.62±11.51)%, (37.37±13.18)%, (38.31±15.66)% in female piglets. With the increase of age, the CD8+ T lymphocytes showed a trend of firstly decrease and then increase. The distribution of lymphocyte subsets in weaned piglets showed different changes with the increase of weaning time, and was influenced by gender. The combination of two different cell delineation methods could reflect more accurate changes of the number and proportion of lymphocyte subsets in piglets.
仔猪在断奶初期, 由于免疫器官尚未完全发育, 对环境的剧烈变化抵抗能力较弱, 很容易感染某些疾病, 造成产量的巨大损失[1]。越来越多的研究表明, T淋巴细胞作为机体免疫系统内功能最重要的一类细胞, 维持着机体正常的免疫功能[2, 3]。在各种临床疾病如自身免疫性疾病、免疫缺陷性疾病、变态反应性疾病、再生障碍性贫血、病毒感染、恶性肿瘤等, T淋巴细胞亚群及其表型都有异常改变。因此, T淋巴细胞亚群(如CD3、CD4 和CD8 T淋巴细胞)的检测对控制疾病的发生、发展, 了解发病机制、指导临床治疗都有极其重要的意义, 是临床检验的一项重要指标[3, 4]。T淋巴细胞亚群的检测一般为检测动物体内免疫状态, 主要是通过检测IFN-γ 及IL-4等细胞因子的含量间接地反映CD4+ 和CD8+ 淋巴细胞的含量, 这种方法不能直接地反映动物体内T淋巴细胞的详细比值, 而流式细胞术作为一种在细胞水平上对单个细胞或其他生物粒子进行定量分析和分选的方法, 现已被广泛应用于检测各种免疫细胞表面标志及细胞内各种细胞因子等[5, 6]。目前, 国内外有关不同日龄仔猪外周血T淋巴细胞亚群的报道较少, 缺乏针对不同生长阶段健康仔猪的免疫系统功能状态的临床研究。本研究利用三色流式细胞术的方法对不同日龄仔猪外周血T淋巴细胞亚群进行测定, 旨在为建立健康仔猪外周血淋巴细胞亚群的临床参考范围提供参考, 比较仔猪在不同生长阶段外周血淋巴细胞亚群的变化趋势, 从而为临床更有效地采用预防治疗措施提供理论依据和临床评价标准。
1.1.1 试验动物
平均体质量8.5 kg且无明显差异(P> 0.05)的“ 杜洛克× 长白× 大白” 三元杂交21日龄断奶仔猪24头, 公母各半, 购自甘肃省兰州市永登县天欣养殖有限公司养殖场。
饲养管理:试验于中国农业科学院兰州畜牧与兽药研究所实验动物基地进行。仔猪饲养于全封闭式猪舍的水泥地面, 饲喂颗粒料, 少量多次, 自由采食、饮水, 不进行后续的免疫及驱虫。
1.1.2 主要试剂及仪器
猪外周血淋巴细胞分离液试剂盒(天津市灏洋生物制品科技有限责任公司)、小鼠抗猪CD3-FITC、小鼠抗猪CD4-SPRD、小鼠抗猪CD8-PE、小鼠IgG-FITC、小鼠IgG-PE、小鼠IgG-SPRD(Southernbiotech公司)、小鼠封闭血清(北京索莱宝科技有限公司)、PBS(上海碧云天生物技术公司)。台式高速冷冻水平离心机、流式细胞仪FC500(美国贝克曼库尔特有限公司)。
1.2.1 仔猪生长速率统计
试验开始后, 分别在断奶后第1、7、21、35天四个时间点的前一天对仔猪进行空腹处理, 次日称量公母仔猪体质量, 统计公母仔猪的生长速率。
1.2.2 断奶仔猪单个核细胞制备
随机挑选24头仔猪, 雌雄各半, 分别在断奶后第1、7、21、35天, 从前腔静脉采集空腹断奶仔猪的外周血, 肝素钠抗凝。按照猪外周血淋巴细胞分离试剂盒说明书分离外周血淋巴细胞。并在收集到的细胞液加入红细胞裂解液, 裂解至澄清透明, 离心弃上清液, D-PBS清洗细胞2遍。采用小鼠封闭血清4 ℃ 孵育30 min, 离心弃上清, D-PBS清洗细胞2遍, 获得断奶仔猪外周血单个核细胞。
1.2.3 流式细胞仪检测方法
(1)同型对照管的制备。配制1× 106 cells· mL-1的单个核细胞, 加入三种同型对照抗体, 4 ℃ 抗体孵育30 min, 离心弃液体, PBS清洗两遍, 500 μ L PBS重悬细胞。
(2)单染管的制备。配制1× 106 cells· mL-1的单个核细胞3管, 分别加入小鼠抗猪CD3-FITC抗体、小鼠抗猪CD4-SPRD抗体、小鼠抗猪CD8-PE标记抗体, 4 ℃抗体孵育30 min, 离心弃液体, PBS清洗2遍, 500 μ L PBS重悬细胞。
(3)样本管的制备。所有样本细胞浓度稀释到1× 106 cells· mL-1, 加入3种小鼠抗猪抗体, 剩余处理同上。
(4)数据分析。采用Beckman FC500流式细胞仪获取细胞, 应用CXP软件进行数据分析, 测得外周血各淋巴细胞的组成及比例。
1.2.4 统计学处理
使用SPSS22.0软件进行统计学分析, 计量资料用Means± SD表示, 组间差异显著性用t检验进行比较, 以P< 0.05为差异有统计学意义。
分别在仔猪断奶后第1、7、21、35天称量仔猪体质量, 统计公母仔猪断奶后体质量, 对结果(图1)进行分析。
通过对公母断奶仔猪体质量数据的分析可知, 公母断奶仔猪在断奶后第1天、第7天体质量差异不显著, 在第21天时公母仔猪体质量分别为(16.02± 2.41)、(18.02± 3.7)kg, 体质量差异显著(P< 0.05), 至断奶后35 d公母仔猪体质量分别为(20.25± 3.48)、(21.98± 3.93)kg, 差异不显著。在断奶后7~21 d, 母仔猪的生长速率大于公仔猪; 在断奶后21~35 d, 公仔猪的生长速率大于母仔猪。
 | 图1 公母仔猪断奶后体质量变化Fig.1 Weight statistics of male and female weaned piglets |
通过分析图2中数据可知, 公母断奶仔猪在断奶开始时CD3+ T淋巴细胞所占外周血单个核细胞的比例分别为(38.00± 3.94)%和(38.17± 3.39)%。仔猪在断奶后CD3+ T淋巴细胞逐渐上升, 至断奶第7天公母断奶仔猪CD3+ T淋巴细胞分别为(40.46± 3.71)%、(41.05± 2.17)%, 差异不显著。至断奶第21 天公母仔猪CD3+ T淋巴细胞亚群比例差异显著, 母仔猪CD3+ T淋巴细胞亚群的比例高于公仔猪, 分别为(47.36± 1.89)%、(42.91± 2.83)%。说明在仔猪生长发育过程中, 母仔猪比公仔猪CD3+ T淋巴细胞较早快速增长, 同时在断奶35日龄时公仔猪CD3+ T淋巴细胞比例较21 d时显著上升, 此时公母仔猪CD3+ T淋巴细胞亚群比例差异不显著。
 | 图2 仔猪外周血CD3+ T淋巴细胞统计(每组样品数为12个; 带有不同小写字母标记的数据间差异显著(P< 0.05))Fig.2 CD3+ T lymphocyte percentage in peripheral blood of piglets(The above values are the average of 12 samples. The values with different letters showed significant difference at P< 0.05 level) |
分析图3中数据可知, 仔猪断奶后0~7 d, 断奶仔猪体内CD4+ T淋巴细胞持续维持在较低水平, 公母仔猪CD4+ T淋巴细胞亚群占外周血单个核细胞的比例差异不显著, 约为10%~11%。至断奶21 d时, 仔猪体内的CD4+ 淋巴细胞才有明显的增长, 所占比例约为14%。至断奶35 d时, 断奶仔猪CD4+ 淋巴细胞亚群已有较高的比例, 达到23%左右, 故随着断奶日龄的增加, 仔猪CD4+ T淋巴细胞亚群的比例逐渐增加。
 | 图3 仔猪外周血CD4+ 淋巴细胞统计Fig.3 CD4+ lymphocyte percentage in peripheral blood of piglets |
分析图4中的数据可知, 在仔猪开始断奶时母仔猪CD8+ 淋巴细胞比例高于公猪, 分别为(28.55± 1.75)%、(23.8± 1.67)%。随着断奶时间的延长, 仔猪断奶7 d时仔猪体内CD8+ 淋巴细胞出现明显下降, 且公仔猪相较于母仔猪下降更明显, 分别为(17.14± 2.64)%、(23.62± 1.51)%。至第21天, 公母仔猪CD8+ T淋巴细胞出现明显上升, 但母仔猪上升的更加明显, 公母仔猪CD8+ T淋巴细胞亚群所占外周血单个核细胞的比例分别为(25.46± 2.32)%、(37.37± 2.18)%。公仔猪至断奶35日龄时, CD8+ T淋巴细胞亚群的比例为(33.33± 2.93)%, 仍低于母仔猪所占比例, 差异不显著, 说明公仔猪CD8+ T淋巴细胞的发育增长稍迟于母仔猪。
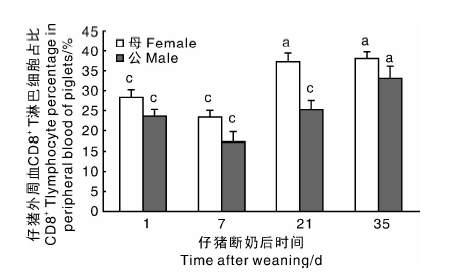 | 图4 仔猪外周血CD8+ 淋巴细胞统计Fig.4 CD8+ lymphocyte percentage in peripheral blood of piglets |
通过分析表1中的数据可知, 仔猪在断奶第1周CD3+ CD4+ T淋巴细胞会出现显著下降, 公母仔猪CD3+ CD4+ T淋巴细胞亚群的比例分别由(6.03± 1.41)%、(5.92± 1.56)%降低至(2.22± 0.24)%、(3.42± 0.85)%, 且这种减少的趋势在外周血单个核细胞及CD3+ T淋巴细胞亚群中是一致的。在外周血单个核细胞群中, 公仔猪CD3+ CD4+ T淋巴细胞下降的比例高于母仔猪, 但在CD3+ T淋巴细胞亚群中公仔猪CD3+ CD4+ T淋巴细胞亚群所占的比例高于母仔猪。21 d时公母仔猪CD3+ CD4+ T淋巴细胞所占比例有所回升, 不同的细胞圈定方法, 会准确地体现出其数量及比例的变化。在母仔猪外周血淋巴细胞中CD3+ CD4+ T淋巴细胞亚群比例高于公仔猪, 但在CD3+ T淋巴细胞亚群中则相反。
通过分析表2中的数据, 随着仔猪断奶时间的增加, 外周血中仔猪CD3+ CD8+ T淋巴细胞数量逐渐增加, 公母断奶仔猪CD3+ CD8+ T淋巴细胞亚群占外周血单个核细胞中的比例由断奶开始时的(4.33± 1.57)%、(5.02± 1.55)%上升到断奶35 d时的(9.59± 2.06)%、(16.54± 2.30)%。其占总外周淋巴细胞的比例逐渐增加, 但其占CD3+ T淋巴细胞的比例在21 d时会有显著的下降, 可能是由于CD3+ T淋巴细胞亚群中其他亚群细胞的增长速率快于CD3+ CD8+ T淋巴细胞。在外周血淋巴细胞中, 公母仔猪从断奶开始时公仔猪CD3+ CD8+ T淋巴细胞亚群的含量持续低于母仔猪。
| 表1 仔猪外周血CD3+ CD4+ T淋巴细胞比例的统计 Table 1 Statistical results of CD3+ CD4+ T lymphocytes percentage in peripheral blood of piglets % |
| 表2 仔猪外周血CD3+ CD8+ T淋巴细胞比例的统计 Table 2 Statistical results of CD3+ CD8+ T lymphocyte percentage in peripheral blood of piglets % |
分析表3中的数据可知, 公母仔猪外周血中CD4+ CD8+ T淋巴细胞随着日龄的增加, CD4+ CD8+ T淋巴细胞所占的比例逐渐增加, 其主要在第21~35天大量升高, 分别由(8.55± 1.19)%、(7.75± 2.41)%上升至(14.44± 3.82)%、(14.10± 3.12)%。但在CD3+ T淋巴细胞亚群中CD4+ CD8+ T淋巴细胞亚群的比例变化则有所不同, 仔猪在断奶第7天的CD4+ CD8+ T淋巴细胞比例较其他日龄显著下降, 差异明显。说明可能是在第7天时CD4+ CD8+ T淋巴细胞增长的速率明显低于其他T淋巴细胞亚群。
| 表3 仔猪外周血CD4+ CD8+ T淋巴细胞比例的统计 Table 3 Statistical results of CD4+ CD8+ T lymphocytes percentage in peripheral blood of piglets % |
通过分析表4中的数据可知, 公母仔猪在断奶初期CD4+ /CD8+ 比值会出现显著性的下降, 分别从断奶开始时的(1.33± 0.23)%、(1.00± 0.26)%, 下降至断奶7 d时的(0.39± 0.25)%、 (0.35± 0.22)%, 这种下降趋势在CD3+ T淋巴细胞亚群中是一致的。至断奶21 d时, CD4+ /CD8+ 会出现显著性升高, 且公仔猪上升的幅度较母仔猪高, 至断奶35 d时公母仔猪比值差异不显著, 此时CD4+ /CD8+ 比值约为0.9。在断奶初期时CD4+ /CD8+ 有较高的比值, 可能是由于在断奶前注射疫苗引起的仔猪免疫细胞CD3+ CD4+ 淋巴细胞的增加, 因而比值较大, 后期随着断奶时间的延长, 且由于断奶应激导致CD3+ CD4+ T淋巴细胞减少。因此, CD4+ CD8+ 比值下降。
| 表4 仔猪外周血CD4+/CD8+值的统计 Table 4 CD4+ /CD8+ values of peripheral blood in piglets % |
由表5可知, 仔猪在断奶开始时有着较高比例的CD4- CD8- T淋巴细胞, 随着断奶天数的增加, 至断奶21 d时逐渐下降, 随后逐渐回升, 但仍然低于开始断奶时的比例。随着断奶仔猪CD3+ 淋巴细胞亚群的增加, 可推测断奶仔猪CD4- CD8- 淋巴细胞在断奶开始至断奶35 d之间其比例变化不大。
| 表5 仔猪CD3+ T淋巴细胞亚群CD4- CD8- T淋巴细胞的比例 Table 5 The proportion of CD4-CD8- T lymphocyte subsets of CD3+ T lymphocyte subsets in piglets unit |
本试验主要是为研究公母仔猪从断奶开始至断奶35 d, 仔猪外周血主要T淋巴细胞亚群的比例变化。T淋巴细胞是机体免疫应答中的重要细胞, 在T淋巴细胞亚群中, CD4+ 细胞数量的减少和功能的低下是仔猪发病的重要因素[7, 8]。由统计数据的结果可知, 随着断奶天数的增加, CD4+ 淋巴细胞数量逐渐增加, 但在断奶后前7 d, 仔猪CD4+ 淋巴细胞持续维持在较低的水平, 且CD4+ 淋巴细胞包含有CD3+ CD4+ T淋巴细胞以及其他CD4+ 淋巴细胞, 本试验结果发现, CD3+ CD4+ T淋巴细胞在断奶7 d时会出现显著性下降, 此时仔猪容易受到外部病原菌的感染, 需要加强日常管理。同时, 在CD3+ T淋巴细胞亚群中, CD3+ CD4+ T淋巴细胞所占比例的变化与外周血淋巴细胞中其所占的比例变化趋势一致, 以上结果说明了在断奶初期由于应激及其他原因仔猪CD3+ CD4+ T淋巴细胞亚群比例出现显著下降。因此, CD4+淋巴细胞的上升可能是由于其他表达CD4分子的淋巴细胞增长所造成的。
CD3+ CD8+ 细胞主要是杀伤性细胞, 含有细胞溶解酶, 用来破坏被感染或受应激的靶细胞; 靶细胞的识别一定程度上是受到刺激, 如应激和感染, 细胞表面表达的非典型MHC分子识别完成的[9]。在对CD8+ 淋巴细胞亚群数据分析时, 发现在仔猪断奶7 d后外周血CD8+ 淋巴细胞的比例会出现显著性下降, 至断奶21 d时CD8+ 淋巴细胞比例较之第7天又会显著上升。仔猪在出生及断奶后逐渐接触到外界环境, 受到外界环境中各种细菌等微生物的影响其CD3+ CD8+ T淋巴细胞逐渐上升, 并且需要维持在一定的比例水平[8]。分析CD3+ CD8+ T淋巴细胞亚群比例时发现, 随着断奶时间的延长呈增长趋势, 同时期公母仔猪CD3+ CD8+ T淋巴细胞比例差异显著, 公仔猪低于母仔猪, 说明在仔猪发育的过程中, 母仔猪CD3+ CD8+ T淋巴细胞的发育先于公仔猪。在断奶仔猪的饲养过程中, 需要特别注意防范公仔猪感染某些病毒病。
T淋巴细胞亚群除CD3+ CD4+ 、CD3+ CD8+ 两大类, 还有CD4+ CD8+ 及CD4- CD8- 两类细胞亚群。通过分析可知, 仔猪在断奶初期, 有着较高比例的CD4- CD8- T淋巴细胞。随着断奶时间的增加, 比例有所降低。但结合CD3+ T淋巴细胞数量增加的趋势, 可推测CD4- CD8- T淋巴细胞占所有外周血单个核细胞中的比例变化不大。同时, CD4+ CD8+ 占所有外周血单个核细胞的比例也逐渐上升, 此类细胞多为记忆性细胞/效应细胞[10, 11], 因此这类细胞对于动物机体抵抗外部病原的再次入侵具有重要作用。在断奶仔猪CD3+ T淋巴细胞亚群中CD4+ CD8+ T淋巴细胞占有很大的一部分, 随着断奶时间的延长, 仔猪CD4+ CD8+ 淋巴细胞在外周血淋巴细胞中所占的比例逐渐增大, 这与猪的CD4+ CD8+ T细胞随着日龄增加而大量增加的结果[12]相一致。CD4+ CD8+ 细胞能够产生IL-10, 而IL-10可以激活B细胞, 使其生长和分化, 并参与抗体的产生, 另外猪CD4+ CD8+ 细胞亚群能产生高水平的IFN-γ [13, 14], 因此对于此类细胞还有待更多的研究。
在T淋巴细胞亚群中, CD3+ CD4+ 细胞数量和功能的低下是仔猪发病的重要因素, CD3+ CD4+ 是CD3+ CD8+ 功能发挥的必需辅助因子, CD3+ CD4+ 与CD3+ CD8+ 合适的比例是维持机体健康的重要因素, 其中CD3+ CD4+ /CD3+ CD8+ 比值的减少是免疫缺陷的重要指征[15]。本试验中通过分析所得结果可知, 在仔猪断奶第7天时, CD4+ /CD8+ 比值低于刚开始断奶时的比值, 说明此时断奶仔猪由于断奶应激导致其免疫细胞数量及比例的变化。在人医临床中, 当CD4+ /CD8+ 比值降至1.5以下时就标志着进入亚健康状态, 并可能成为免疫抑制的重要原因和标志[16, 17, 18], 猪外周血淋巴细胞亚群的组成和比例与其他动物比较有其特殊性。研究表明, 猪外周血淋巴细胞亚群中CD8+ 细胞比例大于CD4+ 细胞, 因此CD4+ /CD8+ 比值也小于人类相应的比值[15]。在仔猪断奶初期CD4+ /CD8+ 大于1, 可能是因为仔猪还未断奶时的免疫程序致使其CD3+ CD4+ 比例高于CD3+ CD8+ 的比例。因此在评价动物的健康状态时仅仅用CD3+ CD4+ /CD3+ CD8+ 的比值是有局限性的, 因此用CD3+ CD4+ 和CD3+ CD8+ 的绝对量及CD4+ /CD8+ 比值作为依据才更加合理。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|