



{kind=link}
{kind=link}
{kind=link}
{kind=link}
溴氰菊酯对中华绒螯蟹肝胰腺氧化胁迫效应和组织结构的影响
[杨宗英1, 2  , 张一柳
, 张一柳1 , 胡鲲1 , 杨先乐1, * , 刘力硕1 , 张凤翔3 , 蔡红桂4 ]
, 张一柳, 刘力硕|
|


作者简介:杨宗英(1988—),女,河南驻马店人,博士研究生,助理工程师,主要从事水产生物免疫学与病害控制研究。E-mail: lxyangzongying@163.com
为探究溴氰菊酯对中华绒螯蟹的毒性和致毒机制,从免疫学和形态学2个方面研究了溴氰菊酯对中华绒螯蟹的毒害作用。采用24 h换水式渔药毒性试验的方法,先测出溴氰菊酯对中华绒螯蟹的24 h-LC50、48 h-LC50、96 h-LC50分别为4.289 4、3.481 8、1.319 5 μg·L-1,安全浓度为0.658 9 μg·L-1;再选取96 h-LC50 的1/2(浓度Ⅰ)、1/5(浓度Ⅱ)、1/10(浓度Ⅲ)3个溴氰菊酯浓度组为试验组,空白组和溶剂组为对照组,分别在给药后测定肝胰腺超氧化物歧化酶(SOD)、过氧化氢酶(CAT)的活性和丙二醛(MDA)积累量等氧化胁迫相关指标的变化;同时在溴氰菊酯胁迫15 d后,观察各浓度组中华绒螯蟹肝胰腺组织结构的变化。结果表明:(1)各个试验组SOD和CAT活力都呈现了先下降后上升再下降的变化趋势,试验组MDA含量始终高于空白组,空白组和溶剂组各个时间点各项指标之间无明显差异。胁迫6 h后,各试验组的CAT的活力极显著低于空白组,其中浓度Ⅱ和Ⅲ组比空白组下降了37.96%和38.92%,MDA含量极显著高于空白组;胁迫12~24 h,SOD和CAT分别受到诱导,24 h时试验组的3个浓度组SOD活力分别比空白组升高了72.13%、87.93%和83.20%,MDA含量减少;胁迫48 h和72 h时,SOD和CAT活力受到抑制,72 h时,浓度Ⅰ和浓度Ⅲ组的SOD分别比空白组下降了40.94%和51.06%,浓度Ⅲ组的CAT值比空白组下降了32.58%,随着胁迫时间的延长,MDA的积累量逐渐增加;(2)在溴氰菊酯胁迫15 d后,各试验组中华绒螯蟹的肝胰腺结构均受到了不同程度的破坏,B细胞数量减少,基膜增厚,肝胰腺上皮细胞肿大并出现空泡样变性,随着溴氰菊酯的浓度增大,肝胰腺受损程度加大,空泡数量增多,上皮细胞脱落,肝胰腺基膜和上皮细胞分离且部分破裂,细胞核固缩。由此得出,溴氰菊酯对淡水中华绒螯蟹属于高毒药物,随着溴氰菊酯胁迫时间的延长和胁迫浓度的增加,肝胰腺MDA含量增加,机体非特异性防御系统受到损伤,肝胰腺正常细胞结构受到破坏。
, ZHANG Yiliu, LIU LishuoIn order to explore the toxicological effects and mechanism of deltamethrin on Eriocheir sinensis, an acute semi-static toxic test was carried out, the LC50 values of 24, 48 and 96 h were 4.289 4, 3.481 8 and 1.319 5 μg·L-1, respectively, and the safe concentration was 0.658 9 μg·L-1. This study also revealed the modulation of antioxidant enzymes such as superoxide dismutase(SOD), catalase(CAT) and malondialdehyde (MDA) content, which was the main product of lipid peroxidation, at 6, 12, 24, 48 and 72 h in different deltamethrin concentration. 96 h-LC50/2(concentration I) group, 96 h-LC50/5(concentration II) group and 96 h-LC50/10(concentration III) group were test groups, and blank group and vehicle group were control groups. Histological structure of hepatopancreas was observed on 15th day in different deltamethrin concentrations. The results were as follows: (1) The oxidative stress indicators were varied in all groups exposed to deltamethrin, SOD and CAT activities showed similar variation of inhibition-induction-inhibition during the test, and the levels of MDA in test groups were all higher than that in blank group. The three indicators had no significant difference at different time in both vehicle group and blank group, and vehicle group also had no obvious difference from blank group. After 6 h exposed to deltamethrin, the CAT activities of all test groups were significantly lower than those of control group, and CAT activities of concentration II and III groups decreased by 37.96% and 38.92% compared with blank groups. MDA levels of test groups were obviously higher than that of control group. After 12 and 24 h, SOD and CAT activities were induced, SOD activities of three test groups were 72.13%, 87.93% and 83.20% higher than that of blank group, and the levels of MDA decreased at 24 h. After 48 h and 72 h, SOD and CAT activities were inhibited again, SOD activities of concentration I and III groups were 40.94% and 51.06% lower than that of blank group, CAT activities of concentration III group were 32.85% lower than that of blank group, and MDA levels of test groups gradually increased along with the increase of stress time of deltamethrin. (2) Histological structure of hepatopancreas was affected at different degrees in different deltamethrin concentration for 15 d. In these three test groups, the number of B cells of hepatopancreas was decreased, the basement membrane became thicker, some hepatic cells were swelling and cavity appeared, some vacuoles appeard in B cells. As deltamethrin concentration increased, the degree of deterioration of hepatopancreas aggravated, the number of vacuoles increased and some hepatic cells were crushing and many cell debris appeared in the lumen, many vacuoles existed between basement membrane and epithelial cells, part of the basement membranes were separated from epithelial cells and ruptured, nucleus were pyknotic. The results indicated that deltamethrin was high toxic pesticide to E.sinensis, with the deltamethrin concentration increasing and stress time prolonging, MDA levels increased, and the non-specific defense system and normal cell structure of E. sinensis were destroyed.
拟除虫菊酯(Pyrethroids)因具有高效、广谱、低毒和能够生物降解等特性得到广泛的应用[1], 溴氰菊酯是拟除虫菊酯类农药中一种重要的广谱杀虫剂[2, 3], 许多地区用于清除野杂鱼和有害生物, 有的甚至用来清塘[4, 5]。拟除虫菊酯对哺乳类和鸟类的毒性不大, 但是对水生动物属于高毒药物, 有研究表明, 拟除虫菊酯对鱼类的毒性比鸟类和哺乳动物高10~1 000倍[6], 目前菊酯类杀虫药对水生生物的研究主要集中在鱼类[7, 8]和虾类[9]。
中华绒螯蟹(Eriocheir sinensis)又称河蟹、毛蟹、清水蟹、大闸蟹, 因其味道鲜美而深受人们喜爱, 是一种重要的淡水养殖动物。我国的中华绒螯蟹养殖模式在不断地改善和发展, 养殖产量也在逐年上升, 统计资料显示, 2009年全国中华绒螯蟹养殖面积达96.62万hm2, 养殖总产值达320亿元[10]。随着中华绒螯蟹养殖产业的发展, 中华绒螯蟹的质量和安全问题也越来越受到人们的关注, 由于农业生产中菊酯类农药的广泛应用以及有些养殖户使用菊酯类农药清塘, 易造成菊酯类农药污染养殖水源, 而对中华绒螯蟹产生毒害作用。有研究指出, 2015年兴化市大面积爆发的中华绒螯蟹“ 水瘪子” 病就是由于溴氰菊酯清塘造成的[11], 目前, 仅见耿雪冰等[12]做了溴氰菊酯对中华绒螯蟹的急性毒性研究, 致毒机理的研究还未见报道。
溴氰菊酯在中华绒螯蟹体内经过细胞色素P450超家族酶类、谷胱甘肽S转移酶和黄素单加氧酶等药物代谢酶的分解会引起活性氧自由基的大量产生[13, 14, 15], 活性氧自由基可以导致蛋白变性、脂质过氧化和DNA损伤[16]。超氧化物歧化酶和过氧化氢酶是甲壳动物抗氧化系统的关键酶, SOD催化
本文在溴氰菊酯对中华绒螯蟹急性毒性试验的基础上, 选择不同的药物浓度对中华绒螯蟹进行毒害胁迫, 研究亚致死浓度下溴氰菊酯对其抗氧化胁迫指标及肝胰腺组织结构的影响, 进而探究中华绒螯蟹“ 水瘪子” 病发病原因, 或可为中华绒螯蟹的健康养殖和疾病防控提供参考。
中华绒螯蟹取自江苏省兴化市安丰镇某养殖场, 个体质量为(114.04± 9.14)g, 在室内暂养一周, 中华绒螯蟹死亡率低于5%且稳定时, 挑选健康、附肢健全、规格相对一致的中华绒螯蟹进行试验。试验用水为充分曝气的居民饮用自来水, 水温为(23.00± 0.05)℃, 水体pH值为7.25± 0.25, 溶解氧为(6.85± 0.15)mg· L-1。试验前1 d停止投喂中华绒螯蟹全价配合饲料。
药品:溴氰菊酯原药(≥ 98%, D9315)购于Sigma公司, 用丙酮配制为10 mg· L-1的母液; 冰醋酸(分析纯), 二甲苯(分析纯), 无水乙醇(分析纯), 苦味酸均购于上海国药集团; 蜡块, 苏木精, 伊红均购于上海生工生物工程有限公司; SOD、CAT、MDA和考马斯亮蓝总蛋白试剂盒均购于南京建成生物工程研究所。
仪器:冷冻离心机(BECKMAN COULTER AgllegraX-15R), 紫外分光光度计(普析通用T6-新世纪), 组织匀浆机(高信化玻), 组织切片机(LEICA RM2135), Olympus显微镜(OLYMPUS BX51)。
1.3.1 溴氰菊酯对中华绒螯蟹的急性毒性试验
选择90 L的水族箱, 放水20 L, 各箱中加入溴氰菊酯的质量浓度分别为0.25、0.5、1、2、4、8、16 μ g· L-1, 进行预试验。正式试验则在致蟹24 h全部死亡的最低浓度和96 h全部存活的最高浓度区间设置6个浓度组, 并以等比数列排布, 浓度系数间隔≤ 2.2[20], 浓度对应为0.5、1、2、4、8、16 μ g· L-1, 同时设置空白组和溶剂组, 溶剂组的丙酮浓度和最高浓度药液中所含丙酮量相同, 另设2个平行试验组, 每个平行试验组放中华绒螯蟹10只, 总共240只试验中华绒螯蟹。试验期间停止喂食, 在试验开始的6 h, 观察各组中华绒螯蟹的中毒状况, 分别在24、48、72、96 h记录各组中华绒螯蟹的死亡数目, 中华绒螯蟹的死亡以鳃停止呼吸为标准, 一旦发现死亡立即清理。
1.3.2 氧化胁迫相关指标的测定
在确定溴氰菊酯对中华绒螯蟹的96 h-LC50 的基础上, 分别设置96 h-LC50 的1/2、1/5、1/10浓度组, 同时设空白组和溶剂组, 各组均设3个平行试验, 每个平行放中华绒螯蟹10只, 总共150只试验中华绒螯蟹, 分别在胁迫6、12、24、48、72 h之后, 从各组中任意取5只中华绒螯蟹, 冰冻麻醉之后, 迅速采取肝胰腺, 用预冷的生理盐水制成10%的匀浆液。
肝胰腺中SOD、CAT和MDA的测定严格按照南京建成试剂盒使用说明书操作。SOD的测定采用黄嘌呤氧化酶法, 定义为每毫克组织蛋白在1 mL反应液中SOD抑制率达到50%时所对应的数值为1个活力单位(U), 单位为U· mg-1 pro; CAT的测定采用240 nm紫外分光光度法, 定义为每克组织蛋白中CAT每秒钟分解吸光度为0.50~0.55的底物中过氧化氢相对量为1个活力单位, 单位为U· mg-1 pro; MDA的测定采用硫代巴比妥酸法, MDA可与硫代巴比妥酸缩合, 形成红色产物, 在 532 nm处有最大吸收峰, 单位为nmol· mg-1 pro; 蛋白含量的测定采用考马斯亮蓝法, 单位为mg pro· mL-1。
1.3.3 肝胰腺组织切片的制备
在96 h-LC50 的1/2、1/5、1/10浓度胁迫第15天, 再分别从3个浓度组和空白对照组随机取出3只蟹, 迅速取出肝胰腺, Bouin氏液固定24 h, 固定样品经70%、80%、95%、100%梯度酒精脱水, 二甲苯透明, 常规石蜡包埋。使用手摇式切片机切片(厚度约5 μ m), H.E(苏木精-伊红)染色, Olympus显微镜观察、拍片。
试验中发现, 刚把中华绒螯蟹放入药液时, 即出现爬箱壁爬气石试图逃出水族箱的行为; 投药4 h后, 中华绒螯蟹开始出现中毒症状, 主要表现出躁动不安, 活动频繁, 身体失去平衡; 随着毒害时间的延长, 中华绒螯蟹开始出现肌肉抽搐, 附肢僵硬、张开, 甚至附肢脱落, 并出现明显的抱团和相互攻击的打架行为。
暴露在不同浓度中的中华绒螯蟹的死亡情况见表1(为3个平行试验的平均结果)。根据TSK[21]可求出24 h-LC50、48 h-LC50、96 h-LC50分别为4.289 4、3.481 8、1.319 5 μ g· L-1, 安全浓度为0.658 9 μ g· L-1。依据GB/T 21281— 2007[23]危险化学品鱼类急性毒性分级试验标准, 可得出溴氰菊酯对中华绒螯蟹属于高毒性, 且随着浓度的增加, 中华绒螯蟹的死亡率升高, 有明显的剂量正相关效应, 空白组和溶剂组中华绒螯蟹的死亡率均为0。
| 表1 不同浓度的溴氰菊酯对中华绒螯蟹的累积致死数 Table 1 Cumulative mortality number of E. sinensis at different deltamethrin concentration |
不同浓度溴氰菊酯胁迫下, 中华绒螯蟹肝胰腺的SOD活力如图1所示, 试验组SOD活力表现出先下降后上升再下降的变化趋势; 在整个试验过程中空白组和溶剂组的SOD活力变化不明显(P> 0.05), 且溶剂组和空白组的差异不显著(P> 0.05), 溴氰菊酯胁迫6 h后, 3个浓度组均出现了SOD活力降低, 但与空白组相比无明显差异; 12 h之后, 3个浓度组的SOD活力出现了迅速上升, 且极显著(P< 0.01)高于空白组, 24 h后, 试验组的SOD活力继续上升, 3个浓度组分别比空白组升高了72.13%、87.93%和83.20%; 48 h后各浓度组SOD活力再次受到抑制, 一直持续到72 h, 各浓度组SOD活力均显著低于空白组, 浓度Ⅰ 组和Ⅲ 组分别比空白组下降了40.94%和51.06%。
 | 图1 溴氰菊酯对中华绒螯蟹肝胰腺SOD酶活力的影响Fig.1 Effects of deltamethrim on SOD activities in hepatopancreas of E. sinensis |
不同浓度溴氰菊菊酯胁迫下, 中华绒螯蟹肝胰腺CAT活力变化见图2, 各时间点溶剂组和空白组的CAT活力无明显变化(P> 0.05), 溶剂组和空白组的CAT活力差异不显著(P> 0.05), 说明溶剂对中华绒螯蟹的肝胰腺CAT活力影响不大。CAT活力出现了与SOD相似的变化规律, 即先下降后上升再下降, 溴氰菊酯胁迫6 h后, 各浓度组CAT活力均出现了极显著(P< 0.01)的下降; 胁迫12 h时, CAT的活力被诱导, 24 h时CAT活力继续升高, 浓度Ⅰ 、浓度Ⅱ 、浓度Ⅲ 组的CAT活力分别比空白组高了56.21%、36.78%和24.74%; 48~72 h, 各浓度组的CAT活力出现持续性降低, 72 h时, 浓度Ⅱ 组CAT活力显著(P< 0.05)低于空白组, 浓度Ⅲ 组极显著(P< 0.01)低于空白组, 比空白组下降了32.58%。
 | 图2 溴氰菊酯对中华绒螯蟹肝胰腺CAT酶活力的影响Fig.2 Effects of deltamethrin on CAT activities in hepatopancreas in E. sinensis |
不同浓度溴氰菊酯胁迫导致的中华绒螯蟹肝胰腺中MDA含量的变化如图3所示, 整个试验过程, 各浓度组的MDA均高于空白组和溶剂组, 溶剂组和空白组在各个时间点的MDA变化不显著, 且溶剂组和空白组的MDA含量差异不明显(P> 0.05), 说明溶剂对中华绒螯蟹肝胰腺中的MDA含量无显著影响。溴氰菊酯胁迫6 h后, 各个浓度组的MDA含量均出现了极显著 (P< 0.01)升高, 到12 h时, 浓度Ⅰ 组、浓度Ⅱ 组和浓度Ⅲ 组分别是空白组的2.77倍、3.20倍和3.25倍; 24 h时, 各浓度组MDA含量出现了明显的降低, 但是在48~72 h, 试验组MDA含量又出现了极显著性(P< 0.01)回升, 肝胰腺处于脂质过氧化状态。
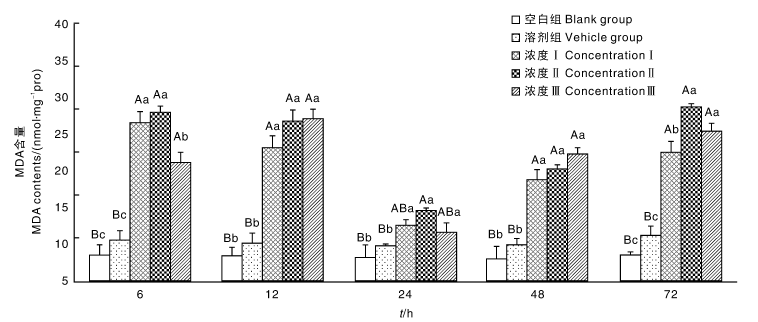 | 图3 溴氰菊酯对中华绒螯蟹肝胰腺MDA含量的影响Fig.3 Effects of deltamethrin on MDA content in hepatopancreas in E. sinensis |
溴氰菊酯不溶于水, 一般用二甲苯为溶剂、农乳2201为乳化剂配制成乳油[24], 本试验用毒性相对较小的丙酮作为溶剂, 急性毒性试验溶剂组中所含丙酮量等同于试验组最高浓度组中含有的丙酮, 结果表明溶剂组和空白组均没有出现中华绒螯蟹死亡, 且溶剂组中华绒螯蟹肝胰腺抗氧化指标和空白对照组的差异不显著, 可见以丙酮配制的溴氰菊酯药液没有显著影响溴氰菊酯的毒性, 溴氰菊酯是关键的致死药物。
组织切片观察发现, 空白组中华绒螯蟹的肝小管结构正常, 上皮细胞排列整齐, 很少有空泡出现(图4), 3个溴氰菊酯浓度组中, 中华绒螯蟹的肝胰腺组织结构分别出现了不同的变化。浓度Ⅲ 组单层柱状上皮细胞肿大, B细胞数量减少, 柱状上皮细胞细胞质中出现空泡, 肝小管基膜增厚; 浓度Ⅱ 组可见上皮细胞中的空泡数量增多, 空泡中出现颗粒物质, 且有部分上皮细胞和管壁脱离, 落入管腔; 浓度Ⅰ 组, 基膜与单层柱状上皮细胞之间形成大量的空泡, 上皮细胞大量脱落, 部分基膜与管壁脱离, 细胞核固缩。
 | 图4 不同浓度的溴氰菊酯对中华绒螯蟹肝胰腺组织结构的影响 Fig.4 Effects of different concentrations of deltamethrin on histological structure of hepatopancreas in E. sinensis |
溴氰菊酯对于哺乳动物和鸟类属于中毒低毒[6], 对鱼类[25]、海洋无脊椎动物[26]、浮游动物[27]以及大型溞[28]属于高毒的研究已有很多报道, 但是菊酯类药物对于甲壳动物的毒性, 则是虾类研究较多, 研究表明, 溴氰菊酯可以影响美国龙虾神经纤维蛋白的磷酸化和脱磷酸化作用[29], 可以使克氏原鳌虾中毒并造成氧化胁迫[30]。但是关于溴氰菊酯对蟹类的毒性研究, 目前国内外仅见耿雪冰等[12]做了溴氰菊酯对中华绒螯蟹的急性毒性研究, 并且溴氰菊酯对中华绒螯蟹成蟹的24 h-LC50、48 h-LC50、96 h-LC50分别为3.16、2.19和0.65 μ g· L-1, 安全浓度为0.46 μ g· L-1, 与本试验的24 h-LC50、48 h-LC50、96 h-LC50分别为4.289 4、3.481 8和1.319 5 μ g· L-1, 安全浓度为0.658 9 μ g· L-1相比更低, 可能是由于中华绒螯蟹规格、养殖环境以及试验条件等不同原因造成的, 但两者的独立试验结果都能说明溴氰菊酯对中华绒螯蟹属于高毒性药物。
甲壳动物具有非特异性免疫, 其主要由血细胞及血细胞被激发释放出的酚氧化酶、溶菌酶和抗氧化酶等多种免疫因子组成[31], 其中SOD和CAT是机体抗氧化系统的关键酶, MDA是脂质过氧化的代谢产物, 这3个指标的变化能反映机体受氧化损伤程度。
本试验中, 在3个溴氰菊酯药液浓度胁迫6 h之后, 各试验组SOD和CAT的活力均出现了显著性降低, 肝胰腺中的MDA含量升高, 说明溴氰菊酯对中华绒螯蟹造成了氧化损伤, 在12 h后, SOD和CAT活力都出现了显著性提高, MDA含量和6 h时变化不大, 说明机体虽然通过增强SOD和CAT的活力对溴氰菊酯的胁迫做出了适应性改变, 但是机体仍然处于脂质过氧化状态; 24 h后, SOD和CAT的活力持续维持高水平, MDA含量也大幅度减少, 与空白组的MDA含量差异性减小, 这说明SOD和CAT的持续高活力, 清除了体内的部分活性氧自由基, MDA含量降低, 中华绒螯蟹的脂质过氧化程度降低; 48 h之后, 中华绒螯蟹肝胰腺中的SOD和CAT活力再次下降, 直至72 h, 浓度Ⅰ 组和浓度Ⅲ 组的SOD活力极显著低于空白组, 浓度Ⅲ 组CAT活力极显著低于空白组, 各试验组MDA含量极显著升高, 这可能是由于溴氰菊酯的长期胁迫产生的大量活性氧超过了SOD和CAT的清除能力, 造成MDA大量积累, 造成细胞损伤。
闫海燕等[32]用氰戊菊酯胁迫鲤后, 鲤肝脏中的SOD表现出了先上升后下降的趋势。本试验中, 溴氰菊酯诱导中华绒螯蟹, 肝胰腺SOD表现出了先下降后上升再下降的趋势, 这可能与不同的药物毒性不同有关, 氰戊菊酯对鲤的96 h-LC50为50.35 μ g· L-1, 而本文计算溴氰菊酯对中华绒螯蟹的96 h-LC50 为1.319 5 μ g· L-1, 毒性差别较大。CAT表现出了同SOD一样抑制-诱导-抑制的变化趋势, 这说明随着溴氰菊酯胁迫时间的延长, 中华绒螯蟹有自身调节相关抗氧化酶的活力, 通过自由基的产生和消除对外界环境做出了适应性改变。
中华绒螯蟹肝胰腺和其他十足目甲壳动物[33]一样, 是甲壳动物重要的消化免疫器官, 主要由4种细胞组成, 分别是B细胞(分泌细胞)、E细胞(胚细胞)、F细胞(吸收细胞)和R细胞(储存细胞)。动物肝胰腺的组织结构变化常常可以反映异物的毒性大小, 毒物的毒性越大, 中华绒螯蟹肝胰腺结构受到破坏的程度越高。
本研究中发现, 在3个浓度的溴氰菊酯药液胁迫下, 中华绒螯蟹的肝胰腺结构受到了不同程度的破坏, 肝胰腺上皮细胞出现大面积的空泡, 并且溴氰菊酯的浓度越高, 中华绒螯蟹的肝胰腺受损越严重, 表现出明显的剂量效应, 这与中华绒螯蟹受到全氟辛烷磺酸盐胁迫[34]和小龙虾受到黄曲霉素胁迫[35]时表现出的症状相似, 推测肝胰腺上皮细胞空泡中出现的颗粒物质可能含有溴氰菊酯或溴氰菊酯的代谢物, 空泡可将这些毒物排除体外, 起到解毒的作用。在溴氰菊酯胁迫下, 中华绒螯蟹肝胰腺基膜的增厚是机体在受到胁迫时产生的胶原纤维和黑色素引起的, 这些物质可抑制病原体的几丁质酶和胞外蛋白酶的活性, 从而抑制或杀死病原体, 是动物的一种自我防御[36]。
除此之外, 甲壳动物肝胰腺中的细胞比例在环境胁迫下会发生变化[37], 不同浓度溴氰菊酯胁迫中华绒螯蟹, 肝胰腺的B细胞减少, 这与中华绒螯蟹受到全氟辛烷磺酸盐胁迫和氨氮胁迫时[38]的结果一致。有研究表明, 甲壳动物B细胞由F细胞分化而来[39], 在F细胞分化为B细胞过程中存在一种P-糖蛋白, 这种蛋白可以消除某些化合物毒性[40], 由此判断中华绒螯蟹在溴氰菊酯胁迫下B细胞减少可能是因中华绒螯蟹利用了肝胰腺中相当数量的B细胞进行解毒, Cheng等[41]在研究中华绒螯蟹对Zn2+的解毒作用中也证明了这一点, 即中华绒螯蟹主要通过肝胰腺的B细胞进行解毒, 当Zn2+浓度超过B细胞的解毒能力时, 会导致中华绒螯蟹肝胰腺损伤, 溴氰菊酯对中华绒螯蟹的毒害是否和重金属的毒害机制完全一致, 还有待进一步验证。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|