




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
大豆异黄酮对雄性大鼠脾脏IL-2、IL-4、TNF-α、INF-γ蛋白表达的影响
[李立科1  , 罗启慧
, 罗启慧1, 2 , 黄超1 , 陈晓林1 , 陈苹1 , 李一帆1 , 刘文涛1, 2 , 陈正礼1, 2, * ]
, 罗启慧]
|
|


作者简介:李立科(1990—),男,四川成都人,硕士研究生,主要从事实验动物疾病模型研究。E-mail:394580507@qq.com
试验旨在探讨不同剂量大豆异黄酮对雄性大鼠脾脏免疫功能的影响及其机理。将40只6周龄雄性SD大鼠随机分为4组,每组10只。对照组和低、中、高剂量组分别灌胃添加不同水平的大豆异黄酮(0、50、250、500 mg·kg-1),大鼠每周称量体质量,连续干预4周后处死大鼠并记录脾脏质量;利用免疫组织化学链霉亲和素-生物素复合物(SABC)染色法研究大鼠脾脏中白介素-2(IL-2)、白介素-4(IL-4)、肿瘤坏死因子α(TNF-α)及干扰素γ(INF-γ)蛋白的表达水平。结果显示,与对照组相比,各组间脾脏指数差异不显著;中、高剂量组脾小体体积显著减小,红髓区淋巴细胞数量显著减少;IL-2在低剂量组表达显著降低,但在中、高剂量组表达增加且在高剂量组达到显著水平;IL-4、TNF-α和INF-γ在大豆异黄酮干预后各剂量组表达水平均显著降低。综上,高剂量的大豆异黄酮能够提高IL-2蛋白的表达水平,各剂量组总体抑制了IL-4、TNF-α和INF-γ蛋白的表达水平,这表明大豆异黄酮对雄性健康大鼠存在潜在的免疫抑制,且呈现一定的剂量差异。
, LUO Qihui
In the present study, effects of different dose of soy isoflavones (SIF) on splenic immune function in male rats were studied and mechanism was explained. Forty rats aged 6 weeks were randomly divided four groups fed with SIF 0 (control group),50 (low dose group),250 (medium dose group), 500 (high dose group) mg·kg-1 body weight. Body weight were recorded every week, and at the end of experiment the rats' spleens were separated and weighed. The expression levels of IL-2, IL-4, TNF-α and INF-γ in rats spleen were detected by immunohistochemistry SABC method. The results showed that compared with control group, there were no significantly differences in spleen index among the groups. Splenic corpuscles in middle and high group were smaller than those in other groups with the lymphocytes number significantly decreased in red pulp. The expression of IL-2 in low dose group was significantly decreased, but rapidly increased in middle and high dose group with a dose-dependent. However, the expression levels of IL-4, TNF-α and INF-γ were all significantly reduced after treated by isoflavones. All above,even though high dose isoflavones can increase IL-2 level,the expressions of IL-4, TNF-α and INF-γ were inhibited with the dose of isoflavones increasing, which indicated SIF had potential immunosuppression to male rats and appeared dosage difference.
大豆异黄酮(Soy isoflavone, SIF)是一种植物雌激素, 同17β 雌二醇具有相似的结构。SIF含有大量的染料木黄酮(genistein)、黄豆苷元(daidzein)和大豆黄素(glycitein)[1], 可作为雌激素α 和β 受体的激动剂或拮抗剂, 从而调节细胞增殖和生长[2]。研究表明, SIF具有雌激素活性和抗雌激素活性双重作用[3], 至于表现为何种活性, 主要取决于其局部浓度、内源性雌激素含量以及组织器官雌激素受体水平[4]。近年来研究表明, SIF具有降血脂[5]、抗氧化[6]、增强免疫[7]等多种生物学功能, 同时, SIF对于癌症、心血管疾病、骨质疏松症和神经退行性疾病也具有良好的预防和治疗作用[8]。脾脏是机体最大的外周免疫器官, 也是免疫细胞定居和产生特异性免疫应答的重要部位[9]。根据细胞因子的分泌方式不同, 可将辅助T淋巴细胞(Th)分为Th1和Th2两个功能亚群, 其中Th1细胞以分泌IL-2、INF-γ 为主, 参与细胞免疫反应; Th2细胞以分泌IL-4、IL-10为主, 参与体液免疫反应[10]。而TNF-α 则由巨噬细胞分泌产生, 可促进细胞增殖和分化。近年来, SIF作为一种保健食品被广泛应用, 但对于其潜在的毒性研究还较少, 如致癌作用[11, 12]、生殖毒性[13]、内分泌紊乱[14]等。因此本试验以雄性大鼠脾脏为研究对象, 探讨补充不同剂量SIF对雄性大鼠脾脏IL-2、IL-4、TNF-α 、INF-γ 蛋白表达的影响, 为揭示SIF对机体免疫功能的作用机理提供理论依据。
1.1.1 试验动物
试验动物为40只6周龄SD雄性大鼠, 体质量(228.44± 11.06)g, 购自成都达硕实验动物有限公司, 使用许可证SCXK(川)2015-030。
1.1.2 试验物品及试剂
试验大鼠专用饲料和垫料均购自成都达硕实验动物有限公司; SIF提取物购自西安天丰生物科技有限公司, 产品批号NF-20140806, 纯度为90%; 兔抗鼠IL-2, IL-4, TNF-α , INF-γ 抗体试剂盒均购自北京博奥森生物技术有限公司; 即用型SABC-AP试剂盒、二氨基联苯胺(DAB)显示试剂均购自武汉博士德生物工程有限公司。
1.1.3 主要仪器
Leica冷冻切片机(德国徕卡有限公司)、Nikon50i-BF荧光生物数码显微镜(日本尼康公司)、江苏捷达801形态分析软件(江苏捷达科技发展有限公司)等。
1.2.1 大豆异黄酮干预试验
将40只大鼠随机分为4组, 在昼夜交替的自然光下饲养, 自由饮水进食。将低、中、高剂量(50、250、500 mg· kg-1)的SIF溶于0.5%羧甲基纤维素钠中, 每日各组大鼠按照2 mL· kg-1 BW连续灌胃4周, 对照组灌胃等剂量的羧甲基纤维素钠, 并每周记录体质量。试验中对动物的处置符合中华人民共和国科技部《关于善待实验动物的指导性意见》的规定。
1.2.2 动物处理
干预4周后, 利用10%水合氯醛腹腔注射麻醉, 断颈处死大鼠。迅速分离出脾脏并称量其质量, 按以下公式计算脾脏指数, 脾脏指数=脾脏质量(mg)/体质量(g), 脾脏置于4%多聚甲醛溶液中进行固定。固定24 h后, 将组织流水冲洗过夜, 随后依次梯度酒精脱水、石蜡包埋、切片(5 μ m)、烤片备用。
1.2.3 H.E染色
利用苏木精-伊红(HE)对切片进行染色, 在光学显微镜下观察脾脏形态结构。
1.2.4 免疫组化染色
将常规石蜡切片浸入二甲苯脱蜡, 经过梯度酒精后, 在新鲜配制的3%过氧化氢溶液中避光反应30 min以清除内源性过氧化氢酶, 然后用PBS冲洗3次, 每次5 min。随后将切片放入柠檬酸钠缓冲液中进行高压抗原修复, 冷却至室温后, PBS润洗3次, 每次5 min。10%山羊血清常温封闭1.5 h后, PBS润洗3次, 随后滴加一抗4 ℃过夜(一抗稀释比例均为1:200), 阴性对照不加一抗。二抗(1:100)常温反应1.5 h后用PBS润洗3次, 每次5 min。滴加链霉亲和素-生物素复合物(SABC)常温反应1 h后, 在光学显微镜下滴加二氨基联苯胺(DAB), 待出现蓝色阳性反应后浸入蒸馏水终止反应, 最后梯度酒精脱水, 二甲苯透明, 中性树胶封片。
1.2.5 图像采集
每组动物10张切片, 对每张切片进行全片统计阳性细胞积分光密度值, 每张切片的积分光密度平均值作为该切片的最终积分光密度值, 10张切片的积分光密度平均值作为该组动物的最终阳性细胞积分光密度值。
利用SPSS 20.0统计软件进行单因素方差分析(one-way ANOVA), 数据以平均值± 标准差表示(
从图1可知, 在SIF干预4周后, 与对照组相比, 大豆异黄酮干预各剂量组脾脏指数均无显著性差异。
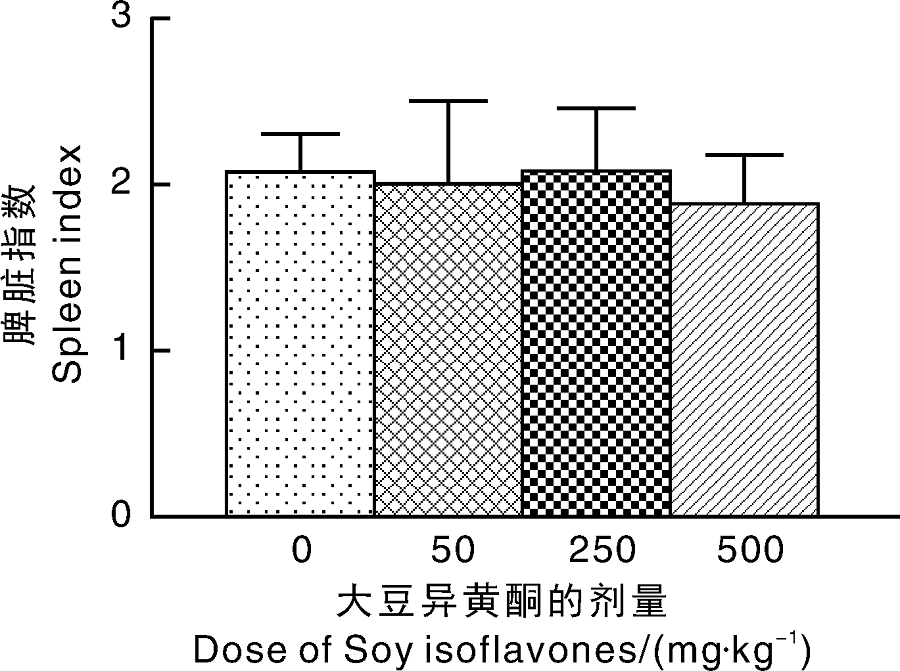 | 图1 大豆异黄酮对大鼠脾脏指数的影响Fig.1 Effect of SIF on spleen index of rats |
如图2所示, 对照组大鼠脾脏组织学结构正常, 脾小体结构清晰, 可见中央动脉; 低剂量组大鼠脾小结直径和红髓区淋巴细胞数量虽有减少, 但与正常组差异不明显; 而中、高剂量组可见脾小体直径显著减小(图3-A), 红髓区淋巴数量显著减少(图3-B), 间质增加。
 | 图2 大鼠脾脏HE染色结果Fig.2 The spleen HE staining of rats |
 | 图3 大鼠脾小结和单位面积淋巴数量变化图中无相同小写字母的数据表示差异显著(P< 0.05), 下同。放大倍数为200× Fig.3 The changes of splenic nodule and number of lymphocyte in spleen of ratsThere is significant difference between the values with no same small letters (P< 0.05); the same below. Magnification 200× . |
SIF对雄性大鼠脾脏IL-2、IL-4、TNF和IFN-γ 蛋白表达影响及积分光密度值显著性统计详见图4、图5。与对照组相比, 低剂量SIF能够显著的降低IL-2蛋白的表达水平(P< 0.05), 而高剂量SIF却能够显著的增加IL-2蛋白的表达水平 (P< 0.05); 组间比较表明低剂量组IL-2蛋白的表达水平显著低于中、高剂量组(P< 0.05, 图4-A)。与对照组相比, 低、中、高剂量SIF组均显著地降低了IL-4、TNF-α 、INF-γ 蛋白的表达水平(P< 0.05, 图4B-D); 组间比较发现中剂量组IL-4蛋白的表达水平显著高于低、高剂量组(P< 0.05, 图4-B), 中剂量组TNF-α 蛋白的表达水平显著高于高剂量组(P< 0.05, 图4-C), 中剂量组INF-γ 蛋白的表达水平显著高于低、高剂量组, 且高剂量组显著高于低剂量组(P< 0.05, 图4-D)。
 | 图4 大豆异黄酮对大鼠脾脏IL-2, IL-4, TNF-α 和INF-γ 蛋白表达的影响Fig.4 Effects of soy isoflavone on IL-2, IL-4, TNF-α and INF-γ protein expression in the spleen of rats |
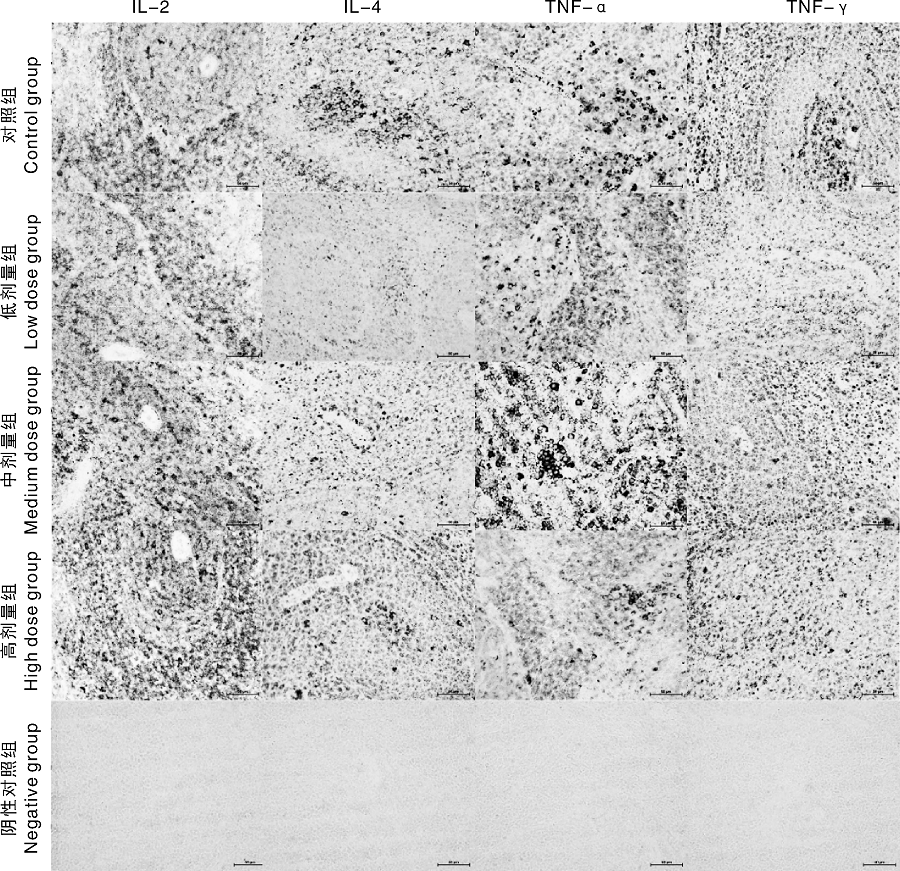 | 图5 IL-2, IL-4, TNF-α 和INF-γ 在大鼠脾脏中的表达(400× )Fig.5 Expression of IL-2, IL-4, TNF-α and INF-γ protein in the spleen of rats (400× ) |
作为亚洲国家的传统饮食, 大豆及其制品中含有丰富的染料木黄酮、黄豆苷元和大豆黄素等成分, 这些异黄酮具有抗氧化、抗炎、调节免疫等功能[15]。许多流行病学研究也表明大豆及其制品能够有效地改善更年期症状、代谢相关疾病, 降低癌症风险[16]。目前大豆食品和异黄酮补充剂的消费量在不断增加, 然而有关SIF的毒性研究发现, 补充异黄酮后会显著增加肿瘤面积[17], 降低内源性雌激素含量, 促进癌细胞生成[11]; 减少雄性动物前列腺重量, 影响睾丸发育, 使雌性动物卵巢异常, 间质萎缩变性[18]等。目前有关SIF对正常雄性大鼠免疫功能的研究报道还较少, 因此对于其作用机制的研究显得非常重要。
脾脏是机体外周最大的免疫器官, 由红髓、白髓和边缘区三个部分组成, 红髓和脾小结主要由B淋巴细胞组成, 白髓的动脉周围淋巴鞘主要由T淋巴细胞组成, 而边缘区则含有大量的巨噬细胞。HE染色结果显示, 中、高剂量组脾小结体积显著减小, 红髓区淋巴细胞数量显著减少。由于机体免疫功能的高低与淋巴细胞的数量密切相关, 而脾小结和红髓含有大量的B淋巴细胞, 在B淋巴细胞显著减少后, 其分化增殖而产生的浆细胞也相应减少, 分泌免疫球蛋白的能力减弱, 最终导致机体体液免疫受到抑制[19]。Th1细胞分泌的IL-2和INF-γ 能够诱发巨噬细胞活化和迟发超敏反应, 进而调节机体细胞免疫[7]。IL-2是引起T细胞增殖的主要细胞因子, 可以刺激NK细胞的生长, 同时增强其杀伤力, 激发B细胞生长及产生抗体。而INF-γ 能够激活巨噬细胞并促进其功能和激活中性粒细胞功能以及NK细胞杀伤力等作用[9]。Th2细胞分泌的IL-4能够诱发肥大细胞和嗜酸性粒细胞生长、分化。巨噬细胞分泌的TNF-α 能够杀伤或抑制肿瘤细胞, 促进细胞增殖和分化, 增强大核细胞的细胞毒作用, 激活LAK细胞和NK细胞。因而IL-2、IL-4、TNF-α 和INF-γ 在机体免疫调节过程中起着重要的作用。免疫组化结果显示, 低剂量的SIF抑制了IL-2 的表达, 然而中、高剂量的SIF却增加了IL-2的表达且在高剂量时达到显著水平。体外研究表明, 在培养的奶牛脾脏和肠系膜淋巴结中添加0.25~5 μ g· mL-1的SIF会增加IL-2的表达, 在培养的鼠脾细胞中添加0.01~10 μ mol的黄豆苷元也会增加IL-2的表达[20]。这表明SIF能够促进IL-2的表达, 而增加的IL-2通过刺激B淋巴细胞的增值和分化, 从而提高B淋巴细胞分泌抗体的功能, 增加机体抗体水平。然而本文发现不同剂量的SIF却显著的抑制了IL-4、TNF-α 和INF-γ 蛋白的表达, 这与大量研究结果相一致, 如Kogiso等[21]给雌性Balb/c小鼠口服4~20 mg· kg-1的染料木黄酮能够显著的减少IL-4和INF-γ 的表达。Davis等[22]给正常健康男性补充50 mg· d-1的SIF会降低TNF-α 的表达。Huang等[23]给绝经后女性口服10~1000 nmol的染料木黄酮和黄豆苷元会降低血清中以及培养单核细胞中的TNF-α 表达水平。Wang[24]和O'Connor[25]分别给卵巢摘除的SD大鼠和雄性Lewis大鼠灌胃和静脉注射20 mg· kg-1的染料木黄酮会减少INF-γ 的分泌。以上表明SIF会影响T细胞和巨噬细胞分泌, 导致IL-4、TNF-α 和INF-γ 蛋白表达减少, 而这些细胞因子表达的减少会抑制细胞的生长、分化和增值, 降低单核细胞和巨噬细胞等杀伤活性, 从而使机体细胞免疫受到抑制。然而也有文献报道异黄酮能够提高免疫功能, 如Zhang等[26]给Swiss小鼠口服黄豆苷元后发现胸腺质量和血液中的淋巴细胞数量显著增加。Guo等[27]研究发现口服2~20 mg· kg-1的染料木黄酮能够增加T细胞和NK细胞活性从而抵抗肿瘤。Sakai等[28]给DO11.10转基因小鼠口服20 mg· kg-1的染料木黄酮后能够显著增加T细胞中IFN-γ 和IL-4的表达。由此可见, SIF是抑制或增强免疫功能, 可能与给药途径、使用剂量、给药时间、动物性别和品种密切相关。
综上所述, 本试验结果显示, 当灌胃剂量超过50 mg· kg-1时, SIF会显著抑制雄性大鼠免疫功能, 这表明补充过量的SIF在雄性大鼠上具有潜在的免疫毒性。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|