

{kind=link}
{kind=link}
SSR荧光标记毛细管电泳法分析30份无花果品种的遗传多样性
[郭正兵1  , 韩柏明
, 韩柏明1 , 郭强2 ]
, 韩柏明|
|


作者简介:郭正兵(1976—),男,江苏高邮人,硕士,副教授,主要从事葡萄、草莓、无花果等果树栽培生理、品源亲缘鉴定等研究。E-mail:260626813@qq.com
为了分析从国内外搜集的无花果品种的遗传多样性,利用SSR荧光标记毛细管电泳检测法构建了30份无花果品种的指纹鉴定平台。以15对SSR荧光引物对30份无花果品种资源进行分析,结果共检测到79个等位变异。15个位点的PIC值在0.332 8~0.875 7,平均为0.530 3;各位点的 Shannon 信息指数(I) 为0.604 9~2.127 3,平均为 1.136 0。30份品种资源的遗传距离在0.025 4~1.123 4。其中引物FCUP038-6的杂合度、PIC 值分别高达0.846 8、0.861 1,可考虑作为以后区试杂交种纯度鉴定的核心标记。30份无花果品种资源表现出比较丰富的遗传多样性,聚类分析结果显示,30份资源被分为两组,来源不同的品种被分散聚类。同时,无花果品种的亲缘关系与其来源和表型特征并不存在明显的相关性。
In order to analyze the genetic diversity of the fig species collected from home and abroad, the capillary electrophoresis detection with fluorescent SSR markers method was used to establish the fingerprint identification platform of 30 fig varieties. Analysis of 30 fig varieties was performed by using 15 pairs of SSR primers. The results showed that a total of 79 alleles were detected, the PIC value of 15 loci was between 0.332 8 and 0.875 7 with an average of 0.530 3 and the Shannon information index (I) of each point was between 0.604 9 to 2.127 3 with an average of 1.136 0. The genetic distance between the 30 varieties was 0.025 4-1.123 4. Among them, the heterozygosity and PIC value of primer FCUP038-6 were up to 0.846 8 and 0.861 1, respectively, which could be considered as the core marker of the purity of the hybrid test. Thirty fig varieties were rich in genetic diversity and were divided into two groups with different origins. There was no significant correlation between the genetic relationships and the origins of the varieties, as well as the phenotype feature.
无花果(Ficus carica L.)属于桑科(Moraceae), 亚热带落叶果树, 多数为小乔木, 也有灌木和大乔木, 是人类驯化最早的经济作物[1]。我国无花果栽培历史悠久, 但一直以来发展缓慢, 只有零星栽培, 直到最近20年来才得到迅速发展, 已经成为一种新兴特色水果。无花果适应性强, 栽培容易, 投资少、见效快、病虫害少, 并具有很高的保健和药用价值, 是小杂果中极具开发前途的果树, 截至2014年底, 据不完全统计, 我国无花果种植面积约为5 000 hm2[2]。因此, 我国无花果产业具有较大的发展潜力。除了新疆、山东栽培较为集中外, 江苏、北京、辽宁等地纷纷开展无花果的引种和试验栽培, 并开展了无花果设施栽培试验。我国无花果栽培品种除了早期引进的新疆早黄、新疆晚黄、青皮、布兰瑞克及紫果等品种外, 自20世纪80年代以来从美国、日本等国相继引入了一些果大、品质优、产量高的新品种, 极大地丰富了我国无花果的品种资源, 并开展了育种研究, 陆续培育了十几个新品种。国外已经开展了较多的关于无花果品种资源方面的研究[3, 4, 5, 6, 7], 但目前我国对无花果种质资源遗传多样性和系谱关系方面的研究还极其缺乏, 相关报道较少[8, 9, 10]。目前在我国无花果生产上, 许多栽培品种名称混淆、来源不清、品种分类工作基础较弱, 使得我国无花果栽培品种无法与世界栽培体系相衔接, 不利于国内无花果种质资源保护与开发利用, 从而造成该树种在新品种选育等方面存在诸多困难[11, 12]。种质资源遗传多样性、亲缘关系和系谱关系的研究是无花果资源的搜集保存和利用的理论基础, 开展这方面的研究是非常必要和迫切的。
DNA分子标记是DNA水平上遗传变异的直接反应, 它们是能稳定遗传的, 而且信息量大, 许多多态性标记在非编码区, 表现选择 “ 中性” , 不受环境因素的影响, 且与基因表达无关, 检测迅速方便快捷, 其在果树分类、品种鉴定、遗传图谱构建和分子辅助选择育种等方面应用广泛, 极大地推动了果树遗传研究[13]。因此, 本文运用荧光SSR技术, 对近年来江苏农林职业技术学院在国内搜集的无花果种质资源进行了遗传多样性方面的初步研究, 旨在为无花果产业的发展提供一些有益参考。
1.2.1 DNA提取
2015年春季取无花果幼嫩叶片作为材料, 进行DNA提取, 利用改进的植物DNA抽提液(添加多种针对植物特点的多糖、多酚去除成分)迅速裂解细胞和灭活细胞内核酸酶, 氯仿抽提后通过离心清除多糖、多酚和蛋白质(根据需要, 上清中还加入异丙醇离心沉淀基因组 DNA, 进一步去除其他各种杂质), 然后在高离子盐状态下基因组 DNA通过选择性吸附于离心柱内硅基质膜中, 经过一系列的快速漂洗、离心等步骤, 进一步去除蛋白、多糖、多酚和细胞代谢物等杂质, 最后用低浓度盐的洗脱缓冲液从硅基质膜上将纯净基因组 DNA洗脱干净(CTAB法植物基因组DNA快速提取试剂盒, 南京钟鼎生物技术有限公司)。提取的DNA用于SSR荧光标记分析。
| 表2 十五对引物序列 Table 2 Sequence of 15 primers |
1.2.2 SSR引物
筛选的15对无花果引物来源于前人研究, 本研究所用荧光引物均由上海生工生物技术有限公司合成, 正向引物加注FAM(蓝) 的荧光染料。
1.2.3 PCR扩增
SSR 荧光引物体系(共 25 μ L):ddH2O 14.8 μ L, dNTP 0.4 μ L, Buffer 2 μ L, 上游引物 0.3 μ L(20 μ mol· L-1), 下游引物 0.3 μ L(20 μ mol· L-1), DNA 模板2 μ L, Taq 0.2 μ L。94 ℃预变性5 min; 94 ℃变性30 s, 54 ℃(退火温度在54 ℃上下波动)复性35 s, 72 ℃延伸40 s, 共35个循环; 最终72 ℃延伸3 min。
1.2.4 毛细管电泳方法
将甲酰胺与分子量内标按100:1的体积比混匀后, 取9 μ L加入上样板中, 再加入1 μ L稀释10倍的PCR产物。然后使用3730XL测序仪进行毛细管电泳, 利用Genemarker 中的Fragment(Plant)片段分析软件对测序仪得到的原始数据进行分析, 将各泳道内分子量内标的位置与各样品峰值的位置做比较分析, 得到片段大小。
一个引物为一个等位基因位点。按照Convert 1.31软件要求的格式录入到EXCEL中, 然后用Convert 1.31软件转化成POPGENE软件所要求格式。使用POPGENE1.32软件进行统计分析。计算等位基因数(A)、观测杂合度(Ho) 、期望杂合度(He)和Shannon信息指数(I) , 采用UPGMA法进行聚类分析。
从60对无花果SSR引物中 选出15对谱带清晰、多态性高的引物。利用选出的15对无花果引物对30个无花果品种材料进行扩增, 扩增产物经过毛细管电泳检测。PIC还是用来衡量某个基因位点等位变异程度高低的指标。试验中15对SSR引物的扩增带型均清晰稳定, 多态性较高且明显, 适合用于无花果SSR 分析。如图1所示, 在30份材料中共检测出79个等位变异, 每对引物检测出2~10个等位变异, 平均每个位点等位基因数为5.27个, 最高为10(FCUP038-6和 LMFC30), 最低为2(MFC8)。15对引物PIC值变化范围在各位点0.332 8~0.875 7, 平均为0.5303。其中最高的为0.846 8(FCUP038-6), 最低的为0.310 3 (LMFC37)。所有引物中PIC含量高于0.5的SSR占到60%。15个位点扩增的片段长度在133~326 bp。观测杂合度(Ho) 0.166 7~0.866 7, 平均为 0.476 8; 期望杂合度(He) 0.332 8~0.875 7, 平均为0.592 9; 各位点的Shannon信息指数(I)0.604 9~2.127 3, 平均为1.136 0。因此, 引物FCUP038-6可作为最佳引物用于鉴定无花果的遗传多样性, 而其他8条引物FCUP044-6、FCUP066-7、FCUP069-6、LMFC28、LMFC30、LMFC38、MFC2、MFC7同时具备较高的杂合率以及多态性, 可考虑作为备选引物。以上结果表明所选取的引物多态性均较高, 有很好的鉴别能力, SSR标记在参试无花果材料中可反映较丰富的遗传多样性信息。
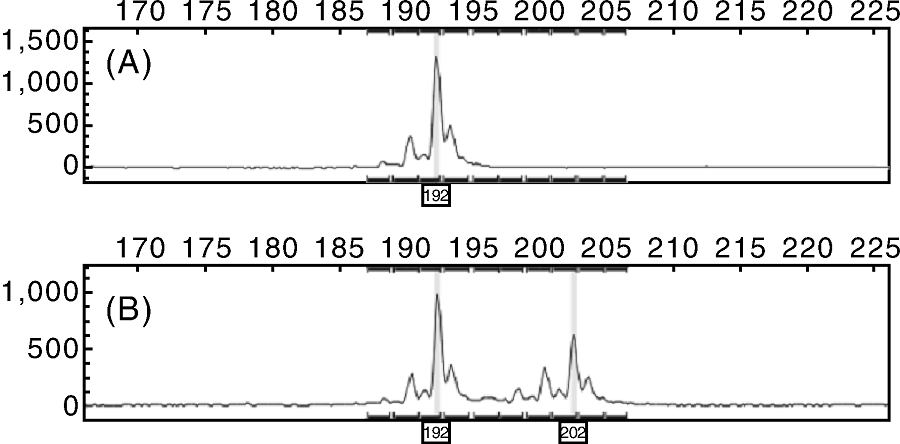 | 图1 华丽(A)和布兰瑞克(B)品种在位点FCUP069-6的电泳图谱Fig.1 The electrophoretic patterns of FCUP069-6 in the gorgeous (A) and Blin Rick (B) cultivars |
经过聚类分析30份无花果品种资源在遗传相似系数0.77处, 可以分别两个类群, 类群Ⅰ 和类群Ⅱ , 在类群Ⅰ 中, 只有3个无花果品种, 分别是华丽、以色列和新疆早黄; 其他27个无花果品种聚为类群Ⅱ 。
| 表3 无花果品种十五对引物的扩增产物及多态性分析 Table 3 Polymorphism analysis of the amplified products of 15 primers |
在类群Ⅱ 中, 在遗传相似系数0.82处又可分为A、B、C三个亚类, 其中A亚类包括布兰瑞克、白热那亚、香蕉、紫色波尔多等19个无花果品种, 其中布兰瑞克和白热那亚被聚在一起; 香蕉、紫色波尔多、加州黑、玛斯义陶芬、亚当、罗伊尔、杜鲁等品种被聚在一起, 其中香蕉和紫色波尔多, 加州黑和玛斯义陶芬, 罗伊尔和杜鲁, 分别被聚在一起, 表现出更紧密的亲缘关系; B1011、波姬红、白马赛、D110、美丽亚、金傲芬、A1213、丰产黄、国王、丽莎等品种被聚在一起, 其中B1011和波姬红, D110和美丽亚, 国王和丽莎, 分别被聚在一起, 表现出更紧密的亲缘关系。在B亚类中只有哈代一个品种。在C亚类中, 包括中国紫果、青皮、紫淘芬、格莱斯、日本紫果、白蜜、紫宝等品种, 其中青皮和紫淘芬, 格莱斯和日本紫果, 分别被聚在一起, 表现出更紧密的亲缘关系。
| 表4 遗传距离与遗传相似系数 Table 4 Genetic distance and genetic similarity coefficient |
 | 图2 基于SSR标记的30份无花果品种聚类图Fig.2 Cluster diagram of 30 Ficus species based on SSR markers |
SSR遗传标记具有试验操作简单、结果稳定、重复性好、共显性稳等特点, 成为研究植物遗传资源多样性的重要工具[13]。研究从60多对无花果SSR引物中筛选出15对稳定性好、多态性高的引物, 采用引物荧光标记技术和毛细管电泳检测技术, 对近年来搜集的30份无花果品种资源开展了遗传多样性分析。试验结果表明, 所搜集的无花果品种资源具有比较丰富的遗传多样性, 这可能与我国无花果品种来源广泛, 是经过长期从不同国家不断引进的结果有关。此外, 无花果品种资源在果实颜色、果实大小、成熟期、开花结果习性以及抗逆性等形态学和生物学方面表现出非常丰富的多样性, 这与本试验中SSR标记的丰富多样性是一致的。
我国不是无花果的原产地, 目前我国栽培的无花果品种资源多数是从国外引进的, 少数是近年自主培育的新品种。无花果在长期的实生繁殖和无性繁殖过程中, 会积累多种多样的表型突变, 这给品种的分类鉴定带来更大的困难。我国多依据无花果的叶片形状和果皮颜色、果形大小等表型进行分类, 经常会发生品种资源的混淆和混乱, 导致同物异名或同名异物。通过单纯依靠形态学进行品种鉴定和分类很可能是不可靠的, 例如, 我国生产上广泛栽培的玛斯义陶芬和波姬红, 叶型和果实形状都非常相似, 常常在生产上被混淆[14, 15, 16]; 本研究的结果证实了这两个品种亲缘关系并不十分密切。在本研究中, 通过SSR聚类分析的结果显示, 青皮和紫淘芬被紧密地聚在一起, 表现出非常近的亲缘关系。但两者果实颜色、大小等形态学差异较大, 青皮果实大小中等, 为绿色, 而紫淘芬果实紫色、较大。而中国紫果、日本紫果和紫色波尔多等紫色品种以及新疆早黄和丰产黄之间并未显示很近的亲缘关系。无花果品种紫淘芬和青皮之间的分化很可能是同一品种由于发生一系列芽变积累的结果, 关于两者之间的亲缘演化关系还有待进一步研究证实[17, 18], 而某些形态学相近的品种其亲缘关系可能很远。在本试验中, 来源不同的无花果品种的聚类是混杂的, 说明无花果品种资源在各国的长期相互引种和选种过程, 导致品种系谱关系十分混乱。本研究初步解析了部分无花果资源间的亲缘关系, 为无花果育种工作提供了初步的理论依据, 对于选育适用于水果市场的鲜食品种, 可优先选择果皮同为紫色、红色、黑棕色的基因型相似度较高的品种作为亲本; 对于选育适用于深加工的品种, 可优先选择22号一族的品种作为亲本。今后还需进一步利用分子标记技术并结合形态学和生物学特性来厘清无花果品种资源的系谱关系。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|