


{kind=link}
{kind=link}
{kind=link}
皂荚种质资源SRAP遗传多样性分析及指纹图谱的构建
[张安世1  , 张素敏
, 张素敏2 , 范定臣3 , 刘莹1 ]
, 张素敏|
|


作者简介:张安世(1965—),男,河南博爱人,硕士,教授,主要从事植物分子生物学研究。E-mail:aszhang1212@163.com
利用SRAP标记技术对18份皂荚种质材料进行了遗传多样性分析。结果表明,从64对SRAP引物中筛选了17对引物进行PCR扩增,共扩增出222个条带,其中多态性条带213个,多态性比率为95.95%。各引物多态性信息含量( PIC)、观测等位基因数( Na)、有效等位基因数( Ne)、Nei's基因多样性指数( H)和Shannon's信息指数( I)的平均值分别为0.861 1、1.962 3、1.415 3、0.257 2和0.406 6,18份皂荚种质资源间遗传相似系数( GS)为0.522 5~0.955 0。UPGMA聚类分析表明,在遗传相似系数为0.66处可将18份皂荚种质资源分为3组,其中,野皂荚单独为1组,山皂荚和皂荚-T聚为1组,其他皂荚材料聚为1组。利用4对引物扩增的9个多态性位点构建了18份皂荚种质资源的DNA指纹图谱,可以将其区分并精准鉴定。该研究结果将为皂荚种质的鉴定、保存和新品种选育提供一定的理论依据。
Genetic diversity of 18 Gleditsia sinensis germplasms were analyzed by SRAP markers. The results showed that 17 primers pairs were screened from 64 primer pairs and 222 bands were obtained, including 213 polymorphic bands, with a polymorphism rate of 95.95%. The average polymorphism information content ( PIC), observed number of alleles ( Na), effective number of alleles ( Ne), Nei's gene diversity ( H) and Shannon's information index ( I) were 0.861 1, 1.962 3, 1.415 3,0.257 2 and 0.406 6, and the genetic similarity coefficients ( GS) among the tested samples ranged from 0.522 5 to 0.955 0. UPGMA analysis showed that 18 Gleditsia sinensis could clustered into 3 groups with the GS of 0.66. Gleditsia heterophylla formed the first group, Gleditsia melanacantha and Zaojia-T were classified into the second group, and the others were classified into the third group. A total of 18 Gleditsia sinensis germplasm DNA fingerprints were constructed from 9 polymorphic loci amplified by 4 primer pairs, and these materials could be distinguished and identified accurately. All these results would provide the important theoretical basis for the identification, conservation and breeding of new cultivar for Gleditsia sinensis germplasms.
皂荚(Gleditsia sinensis Lam.)属豆科苏木亚科皂荚属(Gleditsia Linn.)植物, 为中国特有种, 广泛分布于我国东北、华北、华东、华南等地, 是经济林、用材林、防护林及园林绿化的理想树种[1, 2], 兼有多种优良的生态、经济和社会性能。皂荚还是优良的中药材和工业原料[3], 皂荚、皂刺、皂根、皂叶均可入药, 具有很高的经济价值和药用价值。目前国内外对皂荚的研究主要集中在皂荚果实与皂刺的活性成分[4, 5]、引种栽培[6]、生态属性[2]及遗传学[7, 8, 9]等方面, 分子标记技术也已开始在皂荚研究中应用。邢俊连等[10]利用已公布的皂荚转录组数据设计得到6 494条EST-SSR特异性引物, 并对其中部分引物进行了PCR扩增实验, 结果证实这些引物在其近缘种间具有很高的通用性。李伟等[11]利用AFLP分子标记技术对10个南方皂荚群体进行了遗传多样性研究, 具体分析了皂荚群体遗传结构形成的原因, 并提出了皂荚的保护策略。
SRAP即相关序列多态性扩增(Sequence-related amplification polymorphism), 是由Li等[12]开发的一种新型分子标记技术。该技术针对真核基因中外显子、内含子及启动子的序列差异, 巧妙地设计出2套特异性引物, 可有效地对开放阅读框(open reading frames, ORF)进行PCR扩增。由于被扩增区域为真核基因的ORF, 其本身可能是目的基因的一部分或与目的基因紧密连锁, 因而能对性状进行有效跟踪[13]。SRAP标记在操作上类似于随机扩增多态性DNA标记(random amplified polymorphic DNA, RAPD), 但其稳定性远优于RAPD技术, 同时还克服了SSR技术(simple sequence repeats)标记引物开发的费时费力以及扩增片段长度多态性技术(amplified fragment length polymorphism, AFLP)复杂、成本昂贵等缺点[14], 它提供的信息比其他标记更接近于形态学性状的差异[15]。目前, SRAP标记已广泛应用于植物的遗传多样性分析、品种鉴定、遗传图谱绘制等研究中[16, 17, 18]。
本试验所选用的材料除野皂荚、山皂荚和皂荚-T为实生苗外, 其余的15份材料是以单株产量高、单刺(或单果)平均质量高的植株为接穗、野皂荚为砧木嫁接而获得的无性系, 其中部分品种已经通过河南省林木品种审定委员会审定。另外皂荚-T是从土耳其引进种子的3年生实生苗, 曾被当做皂荚品种, 但其所表现出的形态特征与皂荚有较大差异, 而与山皂荚极为相似。因此, 本试验采用SRAP标记对18份皂荚种质资源进行遗传多样性分析, 并构建DNA指纹图谱, 以期为皂荚种质资源的鉴定、优良品种的选育等提供科学依据。
材料包括3个种共18份, 具体见表1。其中野皂荚、山皂荚和皂荚-T为实生苗, 其余均为嫁接苗。所有材料均随机选取3株, 每株采集2片健康、幼嫩叶片带回实验室置于-80 ℃超低温冰箱保存备用。
| 表1 供试材料 Table 1 Materials used in study |
每个皂荚品种选3株, 每株取1片幼嫩叶片等量混合, 采用改良CTAB法[19]提取皂荚基因组DNA, 并将模板DNA浓度稀释至20 ng· μ L-1, 保存于-20 ℃备用。
选用8个正向引物(Me1-Me8)和8个反向引物(em1-em8)组成64对SRAP引物组合对供试材料进行扩增。反应体积为10 μ L, 包括DNA 1.2 μ L, 上下游引物各0.6 μ L, 2× Taq MasterMix 5.0 μ L, RNase-Free water 2.6 μ L。SRAP-PCR扩增程序为:94 ℃ 5 min; 94 ℃ 1 min, 35 ℃ 1 min, 72 ℃ 1.5 min, 5个循环; 94 ℃ 1 min, 51 ℃ 1 min, 72 ℃ 1.5 min, 35个循环; 72 ℃ 7 min, 4 ℃保存。扩增产物用1.5%琼脂糖凝胶分离。
用64对SRAP引物对供试的18份皂荚材料进行了PCR扩增, 最终筛选出了条带清晰、多态性高的17对引物, 所用引物序列及多态性统计结果见表2。17对引物共扩增出222个条带, 多态性条带213个, 多态性比率的平均值为95.95%。多态性信息含量(PIC)的变化范围为0.799 6~0.919 8, 平均值为0.861 1, 说明所选引物在18个皂荚品种间具有很高的SRAP多态性, 图1为引物组合Me8-em7的扩增结果。各引物的观测等位基因数(Na)的变化范围为1.900 0~2.000 0, 平均值为1.962 3; 有效等位基因数(Ne)的变化范围为1.276 4~1.578 1, 平均值为1.415 3; Nei's基因多样性指数(H)的变化范围为0.193 4~0.329 7, 平均值为0.257 2; Shannon's信息指数(I)的变化范围为0.325 7~0.493 5, 平均值为0.406 6。上述结果表明, 18个皂荚品种间存在较丰富的遗传多样性。同时, 利用SPSS17.0软件对3个主要遗传多样性参数有效等位基因数(Ne)、Nei's基因多样性指数(H)和Shannon's 信息指数(I)进行非参数Kruskal Wallis Test独立样本检验, 结果显示18个皂荚品种间遗传多样性水平存在显著性差异(χ 2=44.308, P< 0.001)。
| 表2 SRAP引物及多态性分析 Table 2 SRAP primers used for this study and their polymorphic |
 | 图1 引物组合Me8-em7的SRAP扩增结果M, 标准分子量; 1~18材料编号同表1Fig.1 The SRAP amplification of primer combination of Me3-em5M, DL 2 000 standard marker; No.1-18 were the same as table 1 |
利用NTSYS-pc软件计算皂荚种质材料间的遗传相似系数(GS), 结果表明, 18份皂荚种质材料间的GS为0.522 5~0.955 0, 平均值为0.727 7, 变幅为0.432 5, 说明供试材料间存在较大的遗传差异。其中焦科4和焦科5的GS最大(0.955 0), 亲缘关系最近, 野皂荚和皂荚-H的GS最小(0.522 5), 亲缘关系最远。
利用NTSYS-pc软件对18份皂荚种质材料进行聚类分析。结果(图2)表明, 在GS为0.66处可将18份皂荚种质材料分为3组。第1组为野皂荚; 第2组为山皂荚和皂荚-T; 第3组共有15份皂荚材料:密刺、嵩刺1、硕刺、皂荚-H、焦科1、焦科3、济科、豫皂1、怀皂王2、博科、豫皂2、太行2、焦科2、焦科4和焦科5。该聚类结果与皂荚的传统分类基本一致。
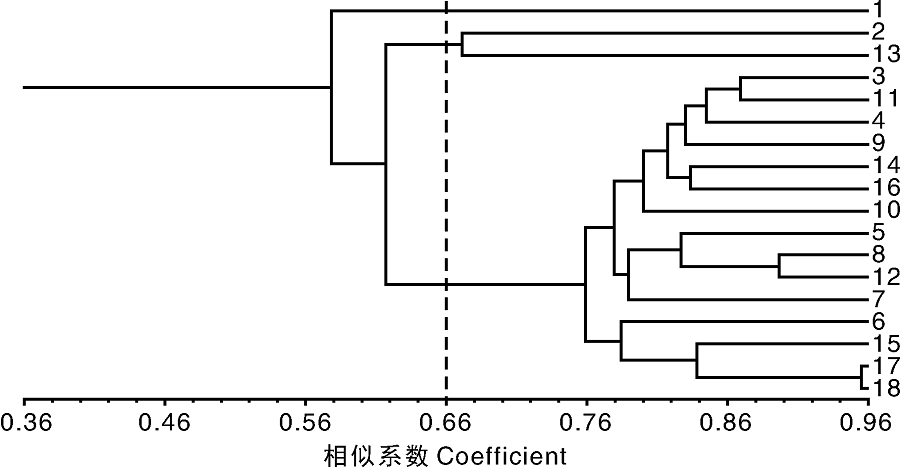 | 图2 十八份皂荚种质资源的聚类图1~18材料编号同表1Fig.2 Dendrogram for 18 G. sinensis germplasm based on SRAP markerNo. 1-18 were the same as table 1 |
通过分析17对引物对18份皂荚种质材料的扩增结果, 选取Me5-em8、Me6-em4、Me8-em7和Me8-em8共4对引物扩增的9个多态性位点构建了18份皂荚种质的DNA指纹图谱(图3)。
 | 图3 十八份皂荚种质资源的DNA指纹图谱1~18材料编号同表1Fig.3 DNA fingerprint of 18 G. sinensis germplasm based on SRAP markerNo. 1-18 were the same as table 1 |
每份材料都有唯一的指纹图谱, 可以将18份皂荚种质材料区分并准确鉴定。
SRAP作为一种新型目的基因分子标记(Gene targeted markers, GTMs), 可以对性状进行跟踪, 被认为是在分子标记辅助育种中最可能被大规模应用的一种技术[22]。与传统的种质资源遗传多样性研究方法相比, 分子标记能直接反映基因组DNA的遗传变异信息, 直接体现该物种在特定环境中遗传多样性水平的高低。因此, 利用SRAP标记技术分析皂荚种质资源的遗传多样性, 了解皂荚种质之间的遗传差异, 对于皂荚种质资源的合理利用及新品种选育具有重要意义。在本研究中, 利用SRAP标记就多个遗传多样性参数分析表明:多态性百分率(PPL)高达95.95%, 多态性信息量(PIC)为0.861 1, 处于高水平(高PIC > 0.5; 0.25< 适中PIC< 0.5; 低PIC< 0.25)[23], 说明本试验筛选的引物具有较高的多态性, 可以有效地用于皂荚的遗传多样性分析。同样, 有效等位基因数(Ne)、Nei's基因多样性(H)和Shannon信息指数(I)分别为1.415 3、0.257 2和0.406 6, 也处于较高水平。另外, 18个皂荚品种间的遗传相似系数(GS)介于0.522 5~0.955 0, 变幅达0.432 5, 说明供试品种间存在较丰富的多样性。对有效等位基因数(Ne)、Nei's基因多样性指数(H)和Shannon's信息指数(I)进行的非参数Kruskal Wallis Test独立样本检验结果表明, 18个皂荚品种间遗传多样性水平存在显著差异, 具有较为丰富的遗传变异。
本研究涉及野皂荚、山皂荚和皂荚3个种共18份材料。聚类结果表明, 在GS为0.66处可将18份皂荚种质材料分为3组。其中, 野皂荚单独为1组, 山皂荚和皂荚-T聚为1组, 其余15份皂荚种质材料聚为1组。从整体结果来看, 该聚类结果与皂荚的传统分类基本吻合。在上述第3组的15份皂荚种质材料中, 焦科4和焦科5是以皂荚荚果为目的选育的品种, 其余均以皂刺为目的选育的品种。聚类结果显示, 焦科4和焦科5在所有供试材料中亲缘关系最近, 与预期相符。从聚类图中还可以看出, 部分品种具有一定的地理来源一致性特征。如采集于河南博爱的5份材料除太行2号外, 其余4份材料豫皂1、豫皂2、怀皂王2和博科聚在一起; 采集于河南新郑的3份材料除皂荚-H外, 山皂荚和皂荚-T聚在一起; 采集于河南修武的5份材料聚为2个分支, 焦科1和焦科3为1个分支, 焦科2、焦科4和焦科5为另一分支。另外, 从聚类结果还获得了一个非常有价值的信息, 即皂荚-T是从土耳其引进种子的3年生播种苗, 起初曾被作为皂荚品种, 但其3年生播种苗无论从叶的形态、皮孔大小、形状还是嫩枝的颜色都与山皂荚极为相似, 因此认为该种子在引进时可能鉴定有误。本研究结果显示, 山皂荚和皂荚-T聚为1组, 两者具有很近的亲缘关系, 因此, 应将皂荚-T归为山皂荚而非皂荚品种, 同时还要结合其他分子标记技术及生长过程中所表现出的其他形态学特征进行进一步的验证。
应用DNA分子标记可直接反映植物品种间的遗传差异, 具有不受环境因子和时空条件影响、高效、快捷等优点, 逐渐取代了依据形态学特征的鉴别方法, 已成为植物品种鉴定的最有效方法。如唐源江等[24]利用17对SRAP引物扩增的48条谱带构建了139份国兰样本的DNA分子身份证, 置信概率达99.99%, 具有唯一性, 可实现对供试国兰品种的精准、快速鉴定。刘君等[25]利用2对SRAP引物构建的指纹图谱, 可以将9个狗牙根品种进行准确鉴别。徐宗大等[26]从100对引物中筛选出了可准确鉴别46个品种的3对SRAP核心引物Me1-Em8、Me2-Em7和Me8-Em9, 构建了玫瑰品种的指纹图谱, 可对46个玫瑰品种和4份野生种质进行准确鉴定。本研究利用SRAP技术, 选用4对引物扩增的9个多态性位点构建了18份皂荚种质材料的DNA指纹图谱, 为皂荚种质资源间的鉴别提供了重要的科学依据。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|