
{kind=link}
{kind=link}
{kind=link}
荞麦ARF基因家族的鉴定及生物信息学分析
[李洪有 , 霍冬敖, 蔡芳, 张晓娜, 石桃雄, 陈其皎, 陈庆富
, 霍冬敖, 蔡芳, 张晓娜, 石桃雄, 陈其皎, 陈庆富* ]
, 霍冬敖, 蔡芳, 张晓娜, 石桃雄, 陈其皎, 陈庆富]
|
|


作者简介:李洪有 (1987—),男,四川资阳人,博士,助理研究员,主要从事植物分子生物技术研究。E-mail: lihongyouluod@163.com
生长素响应因子(auxin response factor, ARF)基因应答了生长素信号,在植物生长发育中具有重要的调控作用。以荞麦基因组数据库为基础,利用BlastP比对程序共鉴定了21个荞麦 ARF基因,并对其基因结构、编码蛋白理化性质、保守结构域、保守基序、亚细胞定位、潜在磷酸化位点及系统进化关系进行了分析。基因结构分析表明,21个荞麦 ARF基因均含有内含子,且不同基因间内含子数目存在较大差异。保守结构域分析显示,21个ARF蛋白均含有保守的B3和ARF结构域,部分ARF蛋白还含有Aux/IAA结构域。蛋白保守基序分析表明,21个ARF蛋白共有10个保守基序,基序长度在13~55个氨基酸之间。亚细胞定位分析表明,大多数ARF蛋白定位于细胞核,个别定位于叶绿体。潜在磷酸化位点分析显示,所有ARF蛋白均含有潜在的丝氨酸(Ser)、苏氨酸(Thr)和酪氨酸(Tyr)磷酸化位点,但各蛋白的不同磷酸化位点的数目差异较大。系统进化分析表明,21个 ARF基因可以分为Ⅰ、Ⅱ、Ⅲ三个亚家族,其中,Ⅰ亚家族可以进一步分为Ⅰa和Ⅰb两个家族,Ⅱ亚家族可以进一步分为Ⅱa和Ⅱb两个家族。研究结果为进一步克隆荞麦 ARF基因及深入研究它们在荞麦中的功能提供了参考。
Auxin response factors (ARF) genes response to auxin signaling and play vital roles in plant growth and development. In the study, a total of 21 ARF genes from buckwheat genome were identified by using BlastP analysis, and then the gene structure, physical and chemical characteristics of encode proteins, conserved domains, conserved motif, subcellular localization, potential phosphorylation sites and evolutionary relationship of these genes were analyzed. Gene structure analysis showed that 21 ARF genes contain intron, and the intron number between genes was different. Conserved domains analysis indicated that all ARF proteins contain B3 and ARF domain, and some proteins also include Aux /IAA domain. Conserved protein motif analysis found that 10 conserved motifs with a range of 13 to 55 amino acids existed in buckwheat ARF proteins. Subcellular localization analysis showed that great majority of ARF proteins located in nucleus and a few of proteins located in chloroplast. Potential phosphorylation sites analysis indicated that all ARF proteins contain Ser, Thr and Tyr phosphorylation sites, but the number of each class’s phosphorylation site between genes was different. Phylogenetic analysis revealed that 21 ARF genes were divided into Ⅰ, Ⅱ and Ⅲ subfamily, and Ⅰ subfamily could be further classed into Ⅰa and Ⅰb subclass, and subfamily Ⅱ could be further divided into Ⅱa and Ⅱb subclass. This study laid a foundation for cloning of buckwheat ARF genes and further functional dissection in growth and development of buckwheat.
生长素信号在植物生长发育过程中起着至关重要的作用。生长素响应因子(auxin response factor, ARF)作为一类重要的转录因子, 其在接受上游传来的生长素信号后, 特异性结合在下游生长素响应基因启动子序列中的生长素响应元件(auxin response elements, AuxRE)TGTCTC上, 激活或者抑制下游靶基因的表达, 进而调控植物的生长发育[1, 2]。研究表明, 所有ARF蛋白都包含一个保守的N-端B3-型DNA结合结构域和一个具有激活或抑制作用的中心ARF功能域。此外, 部分ARF蛋白还含有一个具有二聚作用的C-端Aux /IAA结构域[3]。
目前, 拟南芥基因组中共有23个ARF转录因子基因被鉴定, 研究表明它们调控了拟南芥多个生长发育过程。其中, AtARF1、AtARF2、AtARF3、AtARF4、AtARF5、AtARF7和AtARF19调控了叶发育, 包括对叶长、叶形、叶扩增起始、微管发育、叶脉形成的调控[4, 5, 6, 7, 8]。AtARF2、AtARF3、AtARF4、AtARF5、AtARF7、AtARF8、AtARF10、AtARF16和AtARF19调控了根发育, 包括对胚根形成、侧根形成和生长、根干细胞分化的调控[9, 10, 11, 12, 13]。AtARF1、AtARF2、AtARF6、AtARF8和AtARF17调控了花器官发育, 包括对花粉壁形成、花器官衰老和脱落的调控[14, 15, 16]。此外, AtARF2调控了种子大小, AtARF8调控了角果的形态[17, 18]。目前, ARF基因家族已在包括水稻、玉米、大豆、高粱、谷子、番茄等作物在内的超过30种植物中被鉴定[1, 2, 19, 20]。
荞麦作为一种粮药兼用作物, 具有极高的营养价值和医疗保健作用。目前, 有关荞麦ARF转录因子家族的研究未见报道。鉴定荞麦ARF转录因子并对它们进行生物信息学分析, 对进一步研究它们在荞麦生长发育过程中, 特别是在种子发育过程中的作用具有重要意义。最近甜荞简化基因组测序的完成, 为从全基因组水平上挖掘荞麦ARF家族基因提供了可能[21]。本研究利用BlastP比对程序鉴定了荞麦ARF基因家族, 并对其家族成员的理化性质、亚细胞定位、保守结构域、进化关系、磷酸化位点等进行了预测分析, 为进一步克隆荞麦ARF基因和鉴定它们的生物学功能提供了有用信息。
为了鉴定荞麦基因组中的ARF转录因子, 以23个拟南芥ARF的蛋白序列作为查询序列对荞麦基因组数据库BGBD(Buckwheat Gennome Database, http://buckwheat.kazusa.or.jp/)[20]进行BlastP比对搜索, 序列有E-value< -10确定为初步候选序列。将初步候选序列手动剔除重复序列后进行Pfam保守结构域(http://pfam. xfam. org/)分析[22], 剔除不同时含B3和ARF结构域的蛋白序列, 获得荞麦ARF转录因子家族成员序列。利用BioXM 2.6 软件(http://zhanglab.njau.edu.cn/)计算每个荞麦ARF蛋白的等电点和分子量。
利用WoLF PSORT(http://wolfpsort.org)在线数据库预测每个荞麦ARF蛋白的亚细胞定位情况[23]。
利用HMMER(https://www.ebi.ac.uk/Tools/hmmer/)在线数据库对每个荞麦ARF蛋白的保守结构进行分析[24]。利用MEMM(Multiple Expectation Maximization for Motif Elicitation, http://meme-suite.org/)在线数据库分析每个荞麦ARF蛋白的保守基序, 基序长度设置为6~200个氨基酸, 基序个数设置为10个[25]。
利用NetPhos 3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)在线软件对每个荞麦ARF蛋白的氨基酸序列进行潜在磷酸化位点预测分析[26]。
利用ClustalX 1.83 对21个荞麦和23个拟南芥ARF蛋白序列进行多序列比对, 随后利用MEGA 5.0 软件采用NJ(neighbor-joining)法构建系统发育树, 重复计算值设为1 000次[27]。
通过Blastp比对和Pfam分析, 从甜荞基因组中共鉴定了21个非冗余的ARF基因, 并将它们命名为FeARF1到FeARF21(表1)。21个FeARF基因均含有内含子, 内含子数目为1到14个不等, 其中内含子数目最多的是FeARF9和FeARF10(14个), 最少的是FeARF15和FeARF18(1个)。21个FeARF蛋白长度为294(FeARF8)到1074(FeARF20)个氨基酸不等, 蛋白分子量大小范围为33.46(FeARF8)到118.96 ku(FeARF8)不等。21个FeARF蛋白预测的理论等电点范围为5.38(FeARF10)到9.14(FeARF7), 表明不同的FeARF蛋白可能在不同的微环境中发挥它们的功能(表1)。
| 表1 荞麦FeARF家族基因 Table 1 Predicted FeARF genes in buckwheat |
利用WoLF PSORT对21个FeARF蛋白进行亚细胞定位预测, 结果除FeARF8预测定位于叶绿体和细胞核, FeARF17预测定位于叶绿体外, 其余FeARF蛋白均预测定位于细胞核, 这符合它们作为转录因子调控下游基因表达。
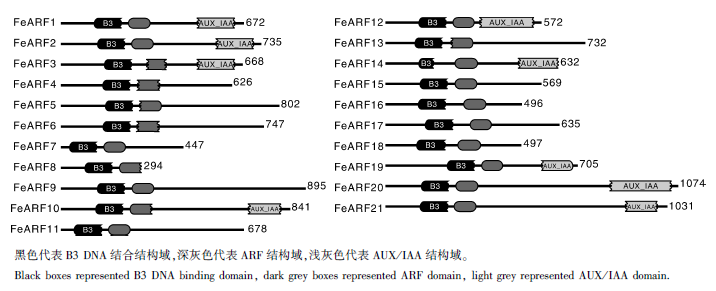
利用HMMER在线数据库对每个荞麦ARF蛋白的保守结构进行分析, 结果如图1所示, 21个荞麦ARF蛋白均含有B3 DNA 结合结构域和ARF结构域。此外, FeARF1、FeARF2、FeARF10、FeARF12、FeARF14、FeARF19、FeARF20和FeARF21还含有Aux/IAA结构域(图1)。
 | 图1 荞麦ARF蛋白的结构域示意图Fig.1 The domain structure of ARF proteins in buckwheat |
利用MEME在线数据库对荞麦ARF蛋白进行保守基序分析, 结果总共10个保守的蛋白基序被鉴定, 每个基序长度在13~55氨基酸之间(表2)。在21个FeARF蛋白中, FeARF1、FeARF2、FeARF5、FeARF6、FeARF9、FeARF10、FeARF12、FeARF19、FeARF20和FeARF21包含了全部10个基序; FeARF3、FeARF4和FeARF13含有8个基序; FeARF14、FeARF15、FeARF16和FeARF17含有7个基序; FeARF7、FeARF11和FeARF18含有6个基序; FeARF8含有5个基序(图2)。利用SMART和Pfam在线数据库对10个基序进行注释发现, 基序1、2代表B3 DNA结合结构域, 基序5、7代表ARF结构域, 基序4、9代表Aux/IAA结构域, 而其他4个基序没有生物学功能被注释, 其生物学功能有待进一步调查。
 | 图2 荞麦ARF蛋白保守性基序分析Fig.2 Conserved motif analysis of buckwheat ARF proteins |
| 表2 荞麦FeARF蛋白保守基序 Table 2 Conserved motif of FeARF proteins |
磷酸化在蛋白激活和信号转导中发挥着至关重要的作用。利用NetPhos 2. 0 Server 软件对21个荞麦ARF家族蛋白的氨基酸序列进行潜在磷酸化位点预测。结果如表3所示, 21个ARF蛋白的氨基酸序列均含有潜在的丝氨酸Ser、苏氨酸Thr和酪氨酸Tyr磷酸化位点, 其中磷酸化位点数目最多是Ser, 其次为Thr(FeARF20除外), 最少的为Tyr。此外, 在21个ARF蛋白中, FeARF20含有最多的磷酸化位点, 共64个; FeARF8磷酸化位点数目最少, 共20个。
| 表3 荞麦ARF蛋白的磷酸化位点预测 Table 3 The predicted phosphorylation sites of ARF proteins from buckwheat |
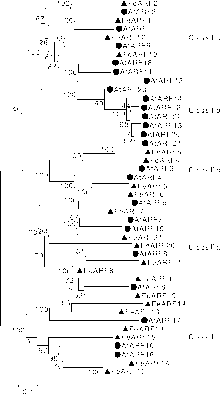
为了进一步理解荞麦ARF基因家族的进化关系, 构建了拟南芥ARF基因家族(23个)和荞麦ARF基因家族(21个)的系统进化树。如图3所示, 44个ARF蛋白可以分为Ⅰ 、Ⅱ 、Ⅲ 三个亚家族, 其中, Ⅰ 亚家族可以进一步分为Ⅰ a和Ⅰ b(只含拟南芥ARF蛋白)两个家族, Ⅱ 亚家族可以进一步分为Ⅱ a和Ⅱ b两个家族。系统进化树分析显示, 除Ⅰ b家族外, 大多数荞麦ARF 蛋白与拟南芥ARF蛋白有“ 一一对应” 关系。此外, 通过系统进化树进一步发现荞麦基因组中存在4对旁系同源基因(FeARF3/FeARF4、FeARF5/FeARF6、FeARF14/FeARF16、FeARF17/FeARF18)。
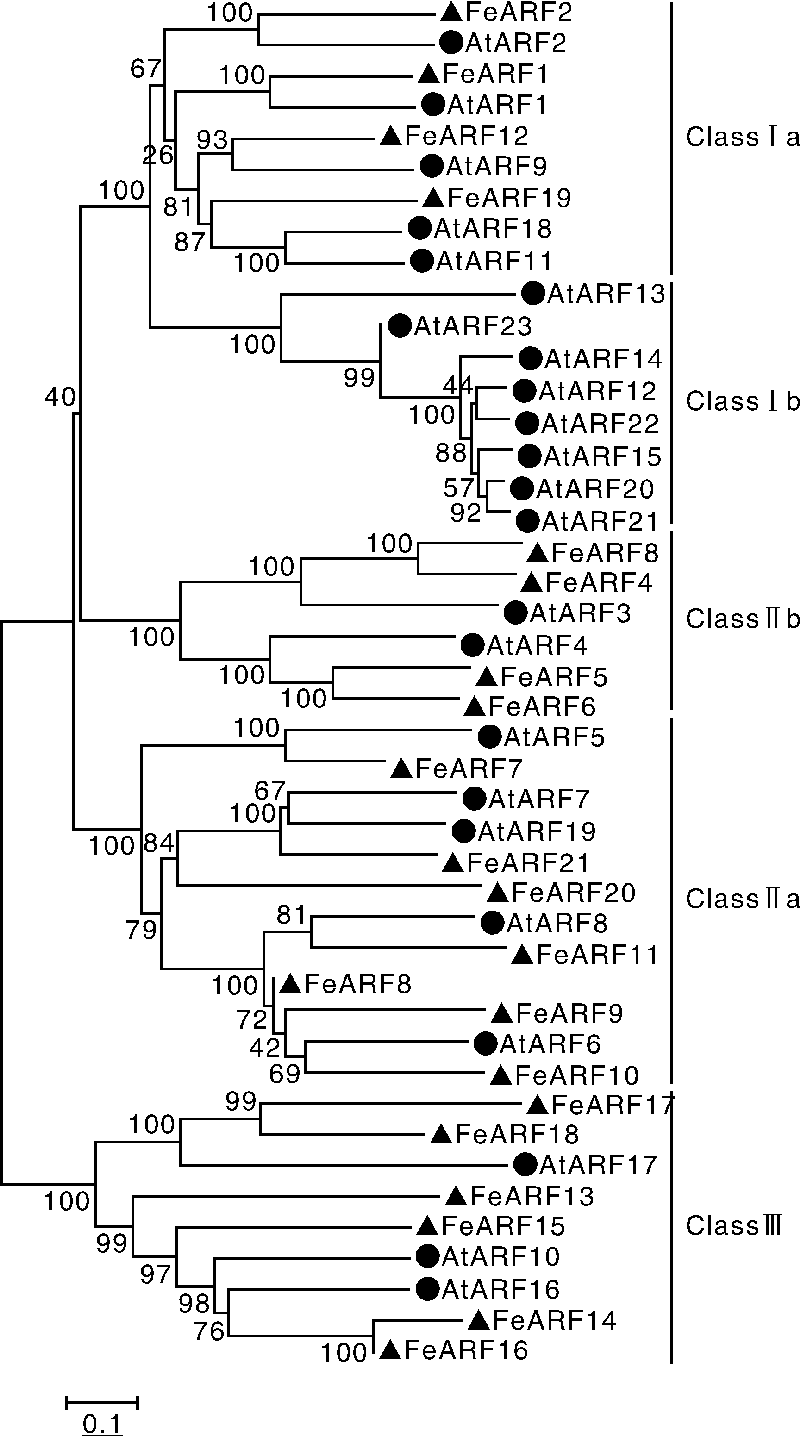 | 图3 荞麦和拟南芥ARF蛋白的进化关系Fig.3 Phylogenetic relationships of ARF proteins from buckwheat and Arabidopsis |
ARF家族基因是生长素信号传导途径中的重要转录因子, 它们接受上游传来的生长素信号, 调控下游一系列生长素响应基因的表达, 在植物生长发育过程中起着至关重要的作用。目前, ARF基因家族已在拟南芥、水稻、玉米、大豆、高粱、谷子、番茄等多种植物中被鉴定[1, 2, 19, 20], 但在荞麦中未见报道。本研究通过生物信息学方法从甜荞基因组数据库中共鉴定到21个荞麦ARF基因, 这与拟南芥(23个)和水稻(25个)ARF基因数目相差不大[28], 表明ARF基因在进化上是保守的。
通常, ARF蛋白具有3个保守的结构域, 即N-端起DNA捆绑作用的B3结构域, 中间具有激活/抑制作用的ARF结构域和C-端具有二聚作用的Aux /IAA结构域[3]。21个荞麦ARF蛋白都含有B3和ARF结构域, 此外, FeARF1、FeARF2、FeARF10、FeARF12、FeARF14、FeARF19、FeARF20和FeARF21还含有Aux /IAA 结构域。这些结果表明, ARF蛋白的蛋白结构在不同植物中是高度保守的。
系统进化树分析表明, 21个荞麦和23个拟南芥ARF蛋白可以分为Ⅰ 、Ⅱ 、Ⅲ 三个亚家族, 其中, Ⅰ 亚家族可以进一步分为Ⅰ a和Ⅰ b两个家族, Ⅱ 亚家族可以进一步分为Ⅱ a和Ⅱ b两个家族。值得注意的是, Ⅰ b家族只包含拟南芥ARF蛋白; 同样, 在多种植物的进化分析中也发现Ⅰ b家族只存在于拟南芥中[1, 2, 19], 表明Ⅰ b家族是拟南芥与其他植物发生分离后在进化过程中新获得的, 可能具有其独特的功能。除Ⅰ b家族外, 大多数荞麦ARF蛋白与拟南芥ARF蛋白具有“ 一一对应关系” , 表明这些蛋白在荞麦和拟南芥发生分离前已存在于二者的共同祖先中, 同时也表明这些蛋白在进化中是高度保守的, 可能具有拟南芥中相似的功能。此外, 通过进化树分析发现了4个荞麦基因复制对(FeARF3/FeARF4、FeARF5/FeARF6、FeARF14/FeARF16、FeARF17/FeARF18), 表明在荞麦和拟南芥发生分离后, 基因复制事件已独立存在于荞麦基因组中。
通过对荞麦基因组中ARF基因家族的鉴定、进化等一系列生物信息学分析, 为将来进一步克隆该家族基因和深入研究该基因家族的功能提供参考, 为揭示该基因家族调控植物生长发育的机制提供理论依据。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|