







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
紫花苜蓿 MsOXO基因的克隆及表达分析
[许青松1, 2  , 赵佳
, 赵佳1, 2 , 魏运民1, 2 , 韩蓉蓉1, 2 , 刘卢生1, 2 , 蒋曹德1, 2 , 玉永雄1, 2, * ]
, 赵佳]
|
|


作者简介:许青松(1994—),女,山东莱芜人,硕士研究生,主要从事牧草遗传育种研究。E-mail:xuqingsong2018@163.com
草酸氧化酶(OXO)在包括核盘菌在内的多种病原体的防御和植物改良中有重大作用。以紫花苜蓿为材料,克隆得到紫花苜蓿草酸氧化酶基因 MsOXO。利用烟草瞬时表达进行亚细胞定位,结果显示,MsOXO定位在细胞壁上。qRT-PCR结果显示, MsOXO在紫花苜蓿的根、茎、叶中均有表达,但在根茎中表达量较低,这可能是根颈和茎基部更易被病菌侵染的原因。外源草酸50 μmol·L-1处理根或100 μmol·L-1处理茎及外源水杨酸100 μmol·L-1处理根均可以诱导 MsOXO基因的上调表达。
, ZHAO Jia
Oxalate oxidase plays an important role in defence of various pathogens such as Sclerotinia and can be used for plant improvement. To explore the role of oxalate oxidase in the defense of Sclerotinia sclerotiorum, alfalfa was used for material in this study, an oxalate oxidase gene, MsOXO, was successfully cloned. Subcellular localization of MsOXO was performed using the transient expression of tobacco, and the result showed that MsOXO was located in cell wall. The results of qRT-PCR showed that MsOXO was expressed in roots, stems and leaves of alfalfa, but was lower in roots and stems, which might be the reason why the root neck and stem base were more susceptible to infection by the pathogen. The up-regulated expression of MsOXO gene was induced by exogenous oxalic acid 50 μmol·L-1 treated root or 100 μmol·L-1 treated stem and exogenous salicylic acid 100 μmol·L-1 treated root.
紫花苜蓿(Medicago sativa)为豆科苜蓿属多年生牧草, 因其优异的饲用价值而被广泛种植, 但病害严重威胁苜蓿的栽培[1, 2, 3]。三叶草核盘菌(Sclerotinia trifoliorum)引起的苜蓿菌核病是世界温带地区广为发生的苜蓿病害[4], 染病植株根茎腐烂、枯萎最终死亡[5]。草酸(oxalic acid, OA)是核盘菌关键致病因子之一[6], 它可以降低细胞间隙pH, 从而增加胞壁降解酶活性并为其他致病因子提供酸性环境[7]; 抑制宿主植物的氧化暴发, 并促进病原体进入[8]; 还可以螯合细胞中的Ca2+, 抑制依赖Ca2+的防卫反应, 阻断Ca2+信号通路[9]; 也能作用于植物表皮细胞, 造成保卫细胞功能紊乱, 改变渗透压, 气孔的不正常开放引起植物叶片萎蔫[10]; 还能激发植物细胞程序性死亡[11]。草酸的降解途径主要是氧化和脱羧。氧化途径是在草酸氧化酶的作用下氧化草酸, 来解除草酸对于寄主的毒害作用, 是一种缓解菌核病害的有效途径。草酸氧化酶(oxalate oxidase, OXO, E.C.1.2.3.4)通过氧化草酸产生H2O2和CO2来解除毒害, 在植物抗病防御和生长发育中起着重要作用[12]。目前, 对OXO基因的研究主要集中在油菜(Brassica napus)和小麦(Triticum aestivum)等作物上。Thompson等[13]和Lane等[14, 15]的研究表明, 小麦萌芽过程中产生的萌发素(germin)是一种具有草酸氧化酶活性的蛋白, 后来陆续在大麦(Hordeum vulgare)、花生(Arachis hypogaea)、油菜、向日葵(Helianthus annuus)等多种植物中发现这种具有草酸氧化酶活性的germin蛋白[16]。Livingstone等[17]在花生中转入大麦OXO基因, 此基因的表达使得花生叶片在感染菌核病后病斑面积减少75%~97%; Thompson等[18]证明表达了OXO基因的转基因油菜对草酸有降解作用, 阻止了由草酸引起的萎蔫。Liu等[19]对过量表达转OXO基因油菜进行研究, 发现其在接种核盘菌后病斑面积和草酸盐含量大幅减少, H2O2水平提高, 综合抗菌核病能力提高。Hu等[20]分析转OXO基因向日葵, 发现OXO基因的表达有效增强了向日葵对菌核病的抵抗能力。王冰山等[21]将小麦OXO基因转入烟草(Nicotiana tabacum), 外源OXO基因的表达提高了转基因烟草离体叶片对草酸的耐受性。胡杰[22]将小麦OXO基因转入拟南芥(Arabidopsis thaliana), 并通过草酸毒素法与菌丝接种法对转基因拟南芥进行了抗病分析, 结果证明转OXO基因拟南芥比野生型拟南芥具有更强的抗病性。目前, OXO基因可以提高抗病性这一功能已在多种植物中得到验证, 而苜蓿中OXO基因却鲜有报道。
本研究通过同源克隆获得紫花苜蓿OXO基因, 利用生物信息学工具对该基因预测分析, 通过亚细胞定位来明确该基因作用区域, 结合qRT-PCR方法对该基因在紫花苜蓿根、茎、叶中的表达水平进行分析, 通过草酸毒素法研究了该基因在各个时间段的表达变化, 外源施用植物激素来研究该基因是否受到诱导表达。本研究旨在通过基因工程的方法提高紫花苜蓿对于菌核病的抗性, 同时也为其他草酸分泌型病原真菌的研究奠定基础。
1.1.1 供试材料
以渝苜一号紫花苜蓿种子为材料, 播种于直径30 cm盛土花盆中, 置于温室棚中进行正常的养护管理。选取健康、长势相近枝条进行扦插水培, 温度20 ° C, 光周期16 h/8 h(L/D)。
1.1.2 主要试剂
RNA提取试剂RNAiso Plus、反转录试剂盒PrimeScriptTM RT reagent Kit with gDNA Eraser (Perfect Real Time)、半定量PCR酶Taq、高保真酶PrimeSTAR、荧光定量PCR酶SYBR Premix Ex Taq Ⅱ 、T4 DNA Ligase、限制性内切酶Sac Ⅰ 、Xba Ⅰ 均购于TaKaRa公司。质粒提取试剂盒Plasmid Mini Kit Ⅰ 、胶回收试剂盒Gel Extraction Kit均购于Omega Bio-Tek公司。大肠埃希菌感受态DH5α 、农杆菌感受态GV3101均购于北京天恩泽生物技术有限公司, 其他试剂为国产分析纯。
1.2.1 RNA提取及反转录
用RNAiso Plus提取紫花苜蓿总RNA, 用1%琼脂糖凝胶电泳检测质量后反转录成cDNA, -20 ℃保存备用。
1.2.2 引物设计、目的基因克隆
根据蒺藜苜蓿OXO基因序列(Medicago truncatula oxalate oxidase, XP_013467999.1)设计引物(OXO-F: 5'-ATGTCCACACAAAAGAACCTTT-3', OXO-R: 5'-CTACCTTTCTTGTTTAGGATCT-3')。内参基因根据紫花苜蓿Actin基因设计(Actin-F: 5'-AAAAGGATGCCTATGTTGGTG-3', Actin-R: 5'-TAAGTGGAGCCTCAGTTAGAAGTA-3')。利用在线荧光定量引物设计工具IDT设计荧光定量PCR引物(OXO-qF: 5'-CTCCCTCAACTTAGCCATCAC-3', OXO-qR: 5'-TGGGAGAATGGTGTGAAAGG-3')。重组载体引物设计为(Sac Ⅰ -OXO-F: 5'-CGAGCTCATGTCCACACAAAAGAACCTTT-3', Xba I-OXO-R: 5'-GCTCTAGACCTTTCTTGTTTAGGATCTAATTT-3')(下划线处为酶切位点)。引物由北京六合华大基因科技有限公司合成。
以紫花苜蓿cDNA为模板, OXO-F和OXO-R为引物, 对目的基因进行PCR扩增。目的片段经电泳纯化回收后连接至pMD19-T载体上, 并转化至大肠埃希菌感受态细胞DH5α , 经菌落PCR筛选后挑取阳性克隆, 摇菌后提取质粒, 将此质粒命名为pMD19T-MsOXO。PCR进一步验证质粒大小, 将验证正确的质粒送往北京六合华大基因科技有限公司测序。
1.2.3 生物信息学分析
使用相关分析工具及数据库对MsOXO基因及其蛋白进行分析, 具体见表1。
| 表1 生物信息学分析工具及数据库 Table 1 Bioinformatics analysis tools and databases |
1.2.4 基因表达分析
不同处理时间下MsOXO基因表达分析:草酸是菌核病菌等草酸分泌型病原菌引起的植物病变的重要致病因子, 核盘菌侵染植株时最先为害根颈及茎基部位, 因此采用草酸毒素法对供试材料的根、茎进行处理。经前期预实验筛选出草酸处理浓度为根部50 μ mol· L-1、茎部100 μ mol· L-1, 此浓度下MsOXO基因正常表达且苜蓿未有明显受害症状。根部处理0、6、12、24、48、72 h后根部取样, 茎部处理0、12、24、48、72 h后茎部取样, 液氮速冻, -80 ℃保存备用。以上处理均重复3次。
不同药物处理下MsOXO基因表达分析:草酸是核盘菌致病因子, 水杨酸(salicylic acid, SA)能够诱导植物抗病性, 是植物防御相关基因表达的激发子, 因此本试验用草酸和水杨酸进行药物试验。水杨酸处理浓度参考前人试验[23, 24]。分别用水、50 μ mol· L-1草酸、100 μ mol· L-1水杨酸处理紫花苜蓿根部。根据不同处理时间下MsOXO在根、茎的表达结果, 确定24 h为取样时间点。根、茎、叶取样, 液氮速冻, -80 ℃保存备用。以上处理均重复3次。
提取以上样品RNA并反转录成cDNA, 以cDNA为模板, OXO-qF、OXO-qR为目的基因引物, Actin-F、Actin-R为内参基因引物, 采用BIO-RAD CFX Connect荧光定量PCR仪进行实时荧光定量PCR。用2-△ △ Ct法处理数据, 分析基因表达水平。用SPSS 19.0软件进行统计分析, GraphPad Prism作图。
1.2.5 重组载体构建及亚细胞定位
以测序正确的pMD19T-OXO质粒为模板, Sac Ⅰ -OXO-F和Xba Ⅰ -OXO-R为引物扩增带有酶切位点目的片段。扩增片段经电泳检测后回收, 连接至pMD19-T载体, 并转化到DH5α 感受态细胞, 经菌落PCR筛选, 阳性菌落经扩大培养后提取质粒, 质粒经PCR进一步验证。用限制性内切酶Sac Ⅰ 和Xba Ⅰ 双酶切上述验证正确的质粒和植物双元表达载体pCambia1300-GFP, 电泳检测后回收, T4连接酶16 ℃过夜连接。连接产物转化DH5α 感受态细胞, 经菌落PCR筛选后测序。将成功构建的重组载体命名为pCam1300-MsOXO-GFP。
提取表达载体pCam1300-MsOXO-GFP质粒, 冻融法转化至根癌农杆菌GV3101感受态细胞, PCR筛选阳性菌株。采用农杆菌介导的烟草瞬时表达进行亚细胞定位, 用注射器将重悬于侵染液中的农杆菌注射至本生烟草(Nicotiana benthamiana)叶片中, 暗培养48 h, 正常培养48 h。撕取侵染后烟草叶片下表皮置于OlympusFV 1000激光共聚焦显微镜下观察、拍照。
扩增所得目的片段与预测大小基本一致, 目的片段经纯化回收连接pMD19-T载体后测序, 结果显示, 该序列全长681 bp, 编码226个氨基酸, 是一个完整的开放阅读框(图1)。BLAST比对分析显示, 该序列与蒺藜苜蓿草酸氧化酶基因MtOXO具有较高的相似性, 推测该基因是紫花苜蓿的OXO基因, 故命名为MsOXO。
 | 图1 MsOXO的核苷酸序列及其编码的氨基酸序列 atg, 起始密码子; -, 终止密码子。Fig.1 Nucleotide and predicted amino acid sequences of MsOXO atg, initiation codon; -, temination codon. |
在线工具ProtParam预测分析表明, MsOXO蛋白分子式为C1117H1753N297O324S9, 分子质量24 815.54, 理论等电点7.06; 在氨基酸组成中, 亮氨酸占比最高(8.8%), 色氨酸及酪氨酸占比最低(0.4%)。该蛋白的正、负电荷的残基均为18; 不稳定系数29.36(< 40), 表明MsOXO为稳定蛋白。
通过SOPMA预测MsOXO蛋白的二级结构, 其中, α -螺旋占比22.12%, 延伸链占比25.22%, β -转角占比7.96%, 无规则卷曲占比44.69%(图2)。经SingalP软件分析, MsOXO蛋白的C(raw cleavage site score)值为0.700, Y(combined cleavage site score)值为0.804, S(signal peptide score)值为0.981, 存在信号肽, 剪切位点在24~25残基处。TMHMM软件预测结果表明, MsOXO蛋白含有1个跨膜区, 其中, 1-6位氨基酸位于细胞膜内侧, 7-24位氨基酸形成一个跨膜螺旋区, 25-226位氨基酸在膜外侧。因此, 信号肽(1-24位氨基酸)被切除后余下的25-226位氨基酸构成的新蛋白质完全位于膜外。
 | 图2 MsOXO蛋白质的二级结构预测 图中线条由高到低依次表示α -螺旋、延伸链、β -转角、无规则卷曲。Fig.2 Secondary structure prediction of MsOXO protein The lines in the figure were alpha helix, extended strand, beta turn and random coil in descending order. |
经TargetP软件分析, MsOXO蛋白的SP(secretory pathway)分值达0.848, 为最高, 表明MsOXO蛋白属于分泌蛋白。采用农杆菌介导的烟草瞬时表达, 在激光共聚焦显微镜下观察MsOXO蛋白, 结果显示绿色荧光集中在细胞壁上(图3)。
 | 图3 MsOXO蛋白亚细胞定位 A, 明场图; B, 荧光图; C, 叠加图。Fig.3 Subcellular localization of MsOXO protein A, Bright field; B, Fluorescent vision; C, merged vision. |
CDD在线分析表明, MsOXO蛋白存在1个保守结构域, 特异性位点为Cupin-1, 属于Cupin超家族, 这一家族都含有由β -折叠构成的桶状结构, Germin是这个家族成员之一。说明MsOXO作为具有草酸氧化酶活性的Germin, 其存在Germin家族的保守结构域。利用PDB预测MsOXO蛋白三级结构, 结果显示, MsOXO高分子实体为草酸氧化酶, 配体为Mn2+, 是一个含锰同型六聚体(图4)。OXO由2个三聚体形成2个结构域, 每个三聚体单体通过相互之间疏水作用和互锁平面保持整个蛋白的稳定性, 每个单体含1个Mn(Ⅱ ), 其与3个His、1个Glu结合, 再与2个分水子配位成球, 构成OXO的活性中心[25, 26]。
 | 图4 紫花苜蓿MsOXO蛋白三维结构预测图Fig.4 Predicted 3D structure of MsOXO protein |
将MsOXO蛋白序列在线BLAST比对得到90多个同源序列, 其与蒺藜苜蓿、鹰嘴豆(Cicer arietinum)、大豆(Glycine max)、木豆(Cajanus cajan)、绿豆(Vigna radiata)、赤豆(Vigna angularis)的OXO基因均有较高相似性, 分别为97%、81%、78%、78%、77%、77%。将以上氨基酸序列经Clustal X及DNAMAN软件进行同源序列比对(图5)。
 | 图5 MsOXO基因编码氨基酸与其他物种相似氨基酸序列比对Fig.5 Homology analysis of amino acid sequence encoded by MsOXO and samilar amino acid sequences from other plant species |
根据同源性分析结果, 选择了20个相似性大于70%的氨基酸序列, 利用MEGA 6.0软件, 采用邻接法(neighbor joining)构建系统进化树(图6)。由进化树可知, 绝大多数相同科的植物聚为一类, 紫花苜蓿、蒺藜苜蓿、大豆、狭叶羽扇豆(Lupinus angustifolius)等同为豆科(Leguminosae)的聚为一类; 青梅(Prunus mume)、山杏(Prunus armeniaca)、甜樱桃(Prunus avium)等蔷薇科(Rosaceae)聚为一类; 长蒴黄麻(Corchorus olitorius)和榴莲(Durio zibethinus)虽属不同科但在聚类分析中聚为一类, 说明MsOXO编码蛋白在同一科属间有很高同源性, 不同科间也有一定的同源性, 推测OXO编码蛋白在进化上较保守。
 | 图6 MsOXO蛋白与其他物种OXO蛋白系统进化树分析2.5 MsOXO基因表达分析Fig.6 Phylogenetic tree analysis of MsOXO protein and OXO protein from other species |
实时荧光定量PCR结果显示:50 μ mol· L-1草酸处理根后, 紫花苜蓿根系中MsOXO表达量在0到72 h内呈先升高后降低趋势, 并在12 h达到最大, 显著(P< 0.05)高于其他时间点表达量, 24 h的表达量次之; 100 μ mol· L-1草酸处理茎后, 紫花苜蓿茎中MsOXO表达量大体呈先升高后降低趋势, 在48 h达到最大, 显著(P< 0.05)高于其他时间点表达量, 说明在外源草酸处理下紫花苜蓿的MsOXO表达量均增加(图7)。
 | 图7 不同浓度草酸处理下MsOXO基因表达情况 A, 50 μ mol· L-1草酸处理苜蓿根部后根中MsOXO表达情况; B, 100 μ mol· L-1草酸处理苜蓿茎部后茎中MsOXO表达情况。不同柱上无相同小写字母表示差异显著(P< 0.05)。下同。Fig.7 Analysis of MsOXO gene expression under different concentrations of OA A, Expression of MsOXO in the root of the alfalfa after 50 μ mol· L-1 OA treatment; B, Expression of MsOXO in the stem of the alfalfa after 100 μ mol· L-1 OA treatment. Data on the bars marked without the same lowercase letter indicated significant differences at P< 0.05. The same as below. |
根据上述不同处理时间下MsOXO表达结果, 为了使根、茎部位MsOXO基因表达量均达到较高水平, 因此在进行不同药物处理试验时选取24 h为取样时间点。试验结果表明, MsOXO基因在无处理的对照紫花苜蓿根、茎、叶中均有表达, 在叶中表达量最多且显著(P< 0.05)高于根茎, 约为根的1.5倍; 经50 μ mol· L-1草酸浸根处理后, 根、茎中MsOXO基因表达量均显著(P< 0.05)升高, 叶中表达量与对照组差异不显著; 经100 μ mol· L-1水杨酸处理后的根、茎、叶中MsOXO表达量较对照均升高, 除根外, 茎叶表达量较对照升高不显著。总体上, 水杨酸处理使得MsOXO基因在根、茎、叶中上调表达(图8)。
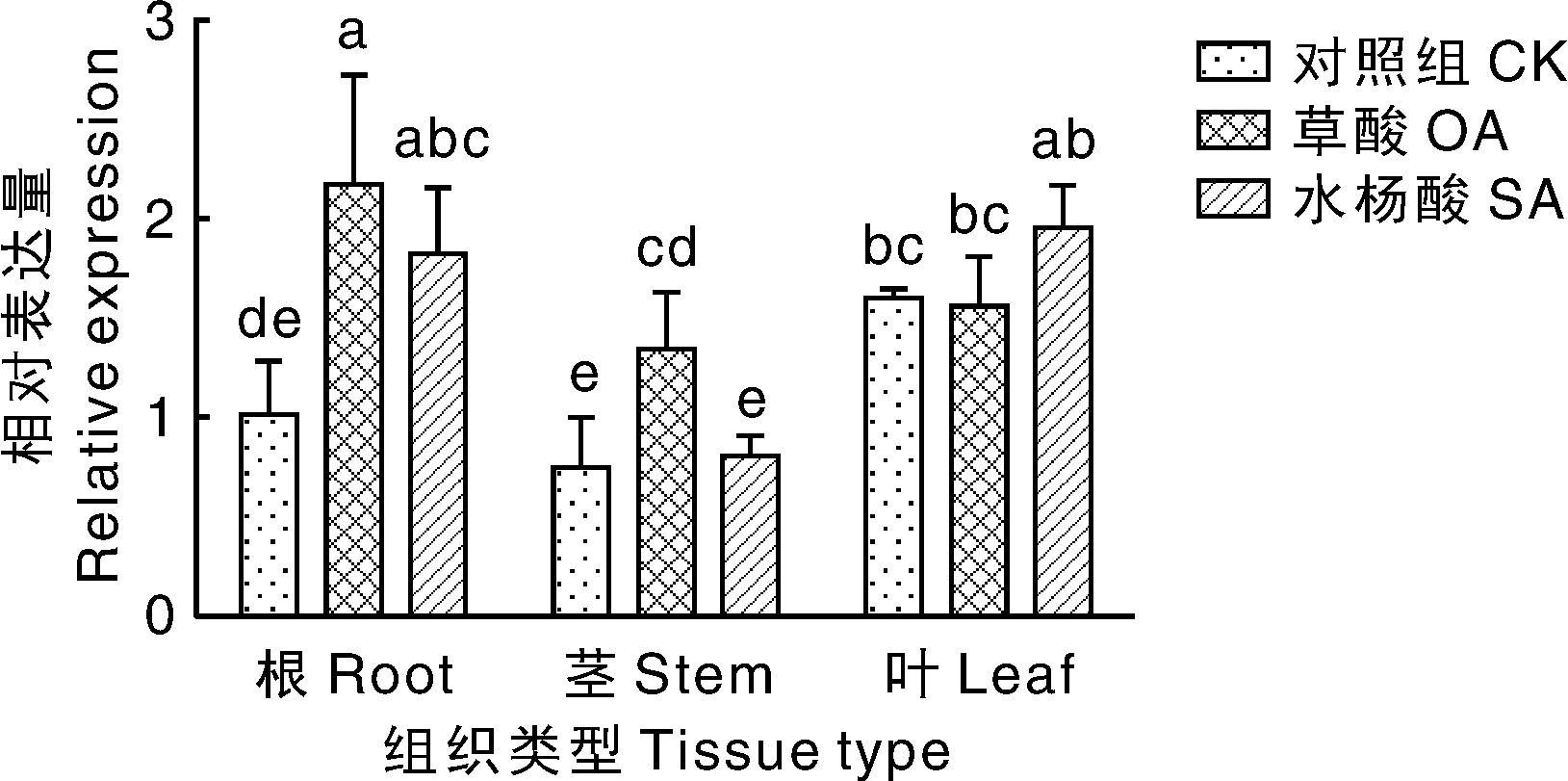 | 图8 草酸、水杨酸处理下MsOXO基因表达情况Fig.8 Expression of MsOXO gene under OA and SA treatments |
本研究通过蒺藜苜蓿OXO基因同源克隆获得了紫花苜蓿草酸氧化酶基因MsOXO。其具有草酸氧化酶的典型结构域, 是一个含锰同型六聚体, 与同属豆科的大豆、鹰嘴豆、羽扇豆等OXO基因有较高的相似度。生物信息学分析结果表明, 其为分泌蛋白, 亚细胞定位结果显示MsOXO蛋白定位于细胞壁, 此结果与前人研究相一致[27]。MsOXO的亚细胞定位结果可为研究其功能及机制提供重要参考信息, 推测MsOXO作为分泌蛋白在胞外解除草酸的毒害, 同时产生H2O2阻止病原真菌进一步损害宿主植物。MsOXO在细胞壁上, 可以在胞外将草酸分解, 防止胞壁水解酶活性上升破坏细胞壁, 同时阻止草酸调控保卫细胞, 也阻止草酸进入细胞危害植物。氧化草酸产生的H2O2发生的氧化作用可以直接在胞外杀死病原菌[30], 还能够引起细胞壁结构蛋白破坏发生融合以阻碍病菌直接进入细胞[28]。病原微生物侵染植物时分泌的草酸造成pH下降, 当pH降到3~5时, 由Glu和Asp构成的带负电氨基酸残基通道打开, 使草酸接近活性中心, OXO分解草酸使pH升高, 升至中性时通道关闭[25]。
植物可以通过诱导一系列防御机制来抵抗病原菌。水杨酸是植物防御相关基因表达的激发子又是重要的内源信号分子, 当植物受到环境胁迫时可诱导一系列防御相关基因的表达而表现出对植物的保护作用。适宜浓度外源水杨酸可以提高苜蓿的抗病性[29]、抗旱性[24]、耐盐性[31]及对低温的抵抗能力[32]。本试验用外源水杨酸处理苜蓿根部, 处理24 h后的组织中MsOXO基因均上调表达, 证明水杨酸对MsOXO基因有诱导表达作用。MsOXO分解病原真菌产生的草酸生成H2O2, 继而产生诱导超氧化物歧化酶(SOD)和过氧化氢酶(POD)等抗氧化酶活性提高, 增强植物对病菌的耐受性[33, 34]。水杨酸作为信号系统介质对H2O2的产生及编码保护蛋白的基因表达有影响[35]。有研究表明, 水杨酸预处理后的小麦种子在感染了颖枯病后, 其叶片组织中编码草酸氧化酶和阴离子过氧化氢酶的基因表达增加[36] 。根据多年田间观察, 西南地区紫花苜蓿菌核病一般在晚秋和(或)早春发生, 且根颈和茎基部最先被侵染, 然后蔓延。本研究中发现, MsOXO在紫花苜蓿的根、茎、叶中均有表达, 在根茎中表达较低, 这也许是紫花苜蓿菌核病首先侵染根茎的原因。以上分析表明, MsOXO基因可能在紫花苜蓿抵御菌核病害中发挥作用。本研究通过对紫花苜蓿草酸氧化酶MsOXO基因的初步分析, 为草酸分泌型真菌病害的研究提供参考。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|