


{kind=link}
{kind=link}
{kind=link}
白羽王鸽卵清蛋白关联蛋白Y基因( OVALY)克隆与生物信息学分析
[徐小钦1, 2, 3  , 杜雪
, 杜雪1 , 李国勤1, 4 , 张龙2 , 陶争荣1 , 卢立志1, 4, * ]
, 杜雪]
|
|


作者简介:徐小钦(1988—),女,四川南充人,博士,研究方向为动物遗传育种。E-mail: xuxiaoqin21@126.com
旨在获得白羽王鸽( Columba livia)卵清蛋白关联蛋白Y基因( OVALY)的cDNA序列,并进行生物信息分析。研究结果显示: OVALY基因cDNA全长为2 068 bp,其中5'非编码区序列180 bp,3'非编码区序列720 bp,开放阅读框1 164 bp,编码的蛋白分子量约44.19 ku,含氨基酸残基388个(GenBank∶KX230792);与NCBI数据库中朱鹮( Nipponianippon) OVALY基因序列同源性最高(81%),与白尾鹰( Haliaeetusalbicilla)和沙鸡( Pteroclesgutturalis) OVALY基因序列的同源性分别为80%和79%。经潜在糖基化位点和磷酸化位点预测分析发现,OVALY蛋白结构中潜在糖基化位点和磷酸化位点分别为4个和22个;经CDD(conserved domain database)数据库分析发现,OVALY蛋白具有丝氨酸蛋白酶抑制剂家族的反应中心区域,属于丝氨酸蛋白酶抑制剂超家族。
The experiment was conducted to clone the whole cDNA of OVALY gene in Columba liviaand analyze the bioinformatic information of the cDNA sequence.The nucleotide sequence of a full-length 2 068 bp cDNA for OVALY was obtained, containing 5'UTR (180 bp), CDS (1 164 bp) and 3'UTR (720 bp), and coding 388 amino acids (44.19 ku, GenBank accession number: KX230792). The OVALY cDNA sequence of white king pigeon was most similar to Nipponianippon, and the homology was 81%. This sequence was also similar to Haliaeetusalbicilla and Pteroclesgutturalis, and the homologies were 80% and 79%, respectively. 4 N-glycosylated sites and 22 phosphorylation sites were detected in these amino acids. Conserved domain analysis of OVALY protein showed this protein had a conserved domain for serpin superfamily.
OVALY(ovalbumin-related protein Y)是蛋清中一种常见蛋白, 在鸡蛋清中含量很少[1, 2], 但在鸽蛋清中却是最主要的蛋白[3]。Gué rin-Dubiard等[4]于2005年采用高通量基因组测序的方式, 获得了鸡的OVALY基因碱基序列, 同时发现该基因存在两种基因亚型。2006年Gué rin-Dubiard等[5]运用双向凝胶电泳技术分离鉴定了鸡OVALY蛋白, 其分子量大小为53 ku。通过软件预测该蛋白分子量为55 ku, 包含21个磷酸化位点和4个N-链接的糖基化位点, 造成此种差异的原因尚未知晓。虽然该蛋白在蛋清中含量少, 但有研究表明在鸡受精卵孵化第7天的卵清中含量迅速增加, 因此推测其可能参与了鸡胚胎发育调控, 具体机制尚不明了[6]。2017年Ren等[3]对鸽蛋清蛋白进行分离, 利用MALDI-TOF-MS技术进行鉴定, 发现OVALY蛋白为鸽蛋清中最主要的蛋白。但关于鸽子OVALY基因及其对应的蛋白的报道鲜有所闻。岩鸽OVALY基因预测序列于2015年已经发布, 但其确切序列尚未获得。
为了研究鸽蛋清中最主要蛋白OVALY的编码基因, 本试验运用RACE技术获得白羽王鸽OVALY基因全部的cDNA序列, 并利用生物信息学数据库和相关软件进行分析, 预测其功能结构。
白羽王鸽输卵管膨大部组织样本采集自平阳县鳌峰鸽业有限公司, 保存于液氮中。
1.2.1 输卵管膨大部中总RNA提取
将适量样本置于盛有液氮的研钵中, 迅速研磨直至粉末状, 加入Trizol试剂, 按照使用说明书进行实验操作, 提取输卵管膨大部总RNA。
1.2.2 反转录
反转录实验试剂采用TransScript First-strand cDNA Synthesis SuperMix(TaKaRa, 大连)试剂盒。严格按照试剂盒的使用说明书进行操作。
1.2.3 OVALY基因克隆
根据岩鸽OVALY(GenBank∶ XM_005509738.1)基因的预测序列, 设计扩增该基因部分片段引物OVALY-F和OVALY-R, 根据获得的OVALY部分片段序列分别设计3'和5'RACE引物, 分别为OVALY-3-GSP1、OVALY-5-GSP1和OVALY-5-GSP2(表1)。引物合成由上海英骏生物技术有限公司完成。
| 表1 OVALY基因引物序列 Table 1 Sequence information of OVALY primers |
根据SMARTTM RACE cDNA Amplification Kit试剂盒(TaKaRa, 大连)说明书步骤扩增5'-UTR和3'-UTR区域; 严格按照pEASY T1 cloning kit试剂盒(TransGen Biotech, 北京)使用说明书进行克隆, 并将获得的产物进行测序[7]。
1.2.4 蛋白质序列分析
利用NCBI的Blast功能进行氨基酸同源性比对。用在线工具ExPASy-PROSITE和ExPASy-ProtParam(http://www.expasy.org)预测蛋白质的基本理化性质。利用SWISSMODEL软件(http://swissmodel.expasy.org)选择同源建模法构建OlCtpE蛋白的三级结构。用NetNGlyc 1.0预测糖基化位点, 用SignalP 4.1预测信号肽。
白羽王鸽OVALY基因cDNA全长为2 068 bp, 其中5'非编码序列为180 bp, 3'非编码序列为720 bp, 开放阅读框为1 164 bp; 编码的蛋白分子量约44.19 ku, 含氨基酸残基388个(GenBank∶ KX230792)。
通过NCBI中Blast功能, 将白羽王鸽的OVALY预测氨基酸序列与其他鸟类的该基因进行比对。发现与朱鹮(Nipponianippon)同源性最高(81%); 与白尾鹰(Haliaeetusalbicilla)、沙鸡(Pteroclesgutturalis)、鸭(Anasplatyrhynchos)、鸡(Gallus)、鹌鹑(Coturnix japonica)的同源性分别为80%、79%、69%、65%、66%。采用N-J的方法构建蛋白质系统发育树, 结果见图1。
 | 图1 系统发育树(N-J)Fig.1 Phylogenetic tree with branch length (N-J) |
利用ExPASy-ProtParam在线软件预测OVALY蛋白的氨基酸构成及性质。该蛋白分子量约44.19 ku, 氨基酸序列中谷氨酸(Glu)和赖氨酸(Lys)的使用频率最高, 分别为10.80%和9.50%, 色氨酸(Trp)使用频率最低(0.8%), 其他氨基酸的使用频率见表2。理论等电点约为5.09, 其中正电荷氨基酸残基49个, 负电荷氨基酸残基63个; 蛋白质氨基酸残基亲疏水性约为-0.315, 脂溶性指数约为85.67, 不稳定系数约为38.00, 体外半衰期约为30 h。说明该蛋白为亲脂的碱性蛋白, 且在体内稳定存在[8, 9]。
| 表2 鸽OVALY氨基酸组成 Table 2 Composition of amino acid of OVALY in pigeon |
通过PSIPRED预测了白羽王鸽OVALY蛋白的二级结构。该蛋白含有10个α 螺旋和14个β 折叠, 由无规则卷曲连接。利用SWISS-MODEL预测鸽OVALY蛋白的三级结构。3D结构模型如图2所示。
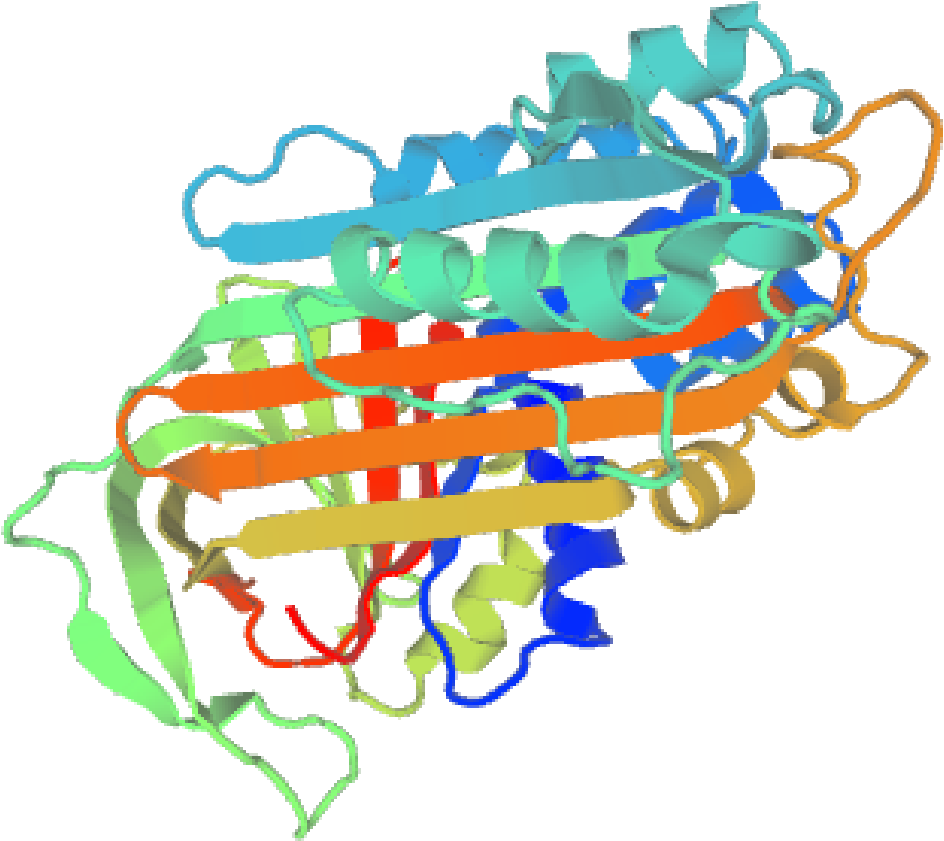 | 图2 鸽OVALY蛋白的3D结构图Fig.2 Tertiary structure of pigeon OVALY protein |
利用NetNGlyc 1.0 Server在线软件预测OVALY蛋白序列中Asn-Xaa-Ser/Thr三肽概率大于50%的N-连接糖基化位点, 共发现了4个, 分别位于62、95、215和374处; 利用NetOGlyc 3.1 Server在线软件未发现概率大于50%的O-GalNAc连接糖基化位点。利用NetPhoQ2.0server在线软件预测概率大于50%的磷酸化位点, 共发现了22个, 其中丝氨酸(Ser)15个, 分别位于34、52、83、97、99、148、200、261、308、314、318、321、345、355和356处; 酪氨酸(Tyr)5个, 分别位于16、79、107、235和385处; 苏氨酸(Thr)2个, 分别位于202和343处。
通过CDD数据库对OVALY蛋白保守结构域进行分析, 发现该蛋白具有丝氨酸蛋白酶抑制剂反应中心结构域, 属于丝氨酸蛋白酶抑制剂超家族成员(图3)。
 | 图3 鸽OVALY蛋白的结构域分析Fig.3 Conserved domain analysis of pigeon OVALY |
SignalP4.0在线软件分析, 在该蛋白中不具有信号肽区域; 利用TMHMM Server v. 2.0软件分析, 该蛋白不具有跨膜区域。因此, 该蛋白可能通过非经典分泌途径完成分泌。
基因翻译成氨基酸序列后, 主要以糖基化和磷酸化进行修饰, 在许多生物进程中具有重要作用。研究表明, 蛋白的糖基化参与蛋白质折叠、细胞识别、免疫反应等; 磷酸化在信号转导、基因表达、细胞周期调控中起到关键性作用[10, 11, 12, 13, 14]。具有免疫功能的蛋白大多都含有糖基化修饰位点, 在生物医学研究方面具有重要意义[15]。在白羽王鸽和鸡的OVALY蛋白中均含有4个N-连接糖基化位点, 磷酸化位点分别为22个和21个, 两个物种的该蛋白具有相似的翻译后修饰。蛋白质的翻译后修饰, 对蛋白质的结构和功能具有重要作用, 因此对被修饰的结构位点进行深入分析具有重要意义。
真核蛋白质分泌包括经典和非经典两种分泌途径。经典分泌途径是指内质网-高尔基体途径, 这种途径是大多数蛋白的分泌方式; 只有小部分无信号肤的蛋白质不依赖该途径, 就能从胞内运输到细胞外, 因此叫做非经典分泌蛋白。一般非经典分泌蛋白是相对分子量12~45 ku的小分子蛋白, 缺少潜在糖基化位点和信号肽等结构, 受温度、能量等影响较大[16]。本研究中白羽王鸽OVALY蛋白序列中不具有信号肽区域和跨膜结构, 且分子量大于12 ku, 小于45 ku, 因此推断该蛋白的分泌方式为蛋白质非经典分泌途径; 但是该蛋白又包含了4个潜在糖基化和22个潜在磷酸化位点, 故而其分泌方式有待进一步探讨[17]。
经蛋白质保守结构域分析发现, 鸽OVALY蛋白具有丝氨酸蛋白酶抑制剂超家族(Serpin superfamily)特有的反应活性中心, 属于丝氨酸蛋白酶抑制剂超家族成员。在鸡蛋中共发现了15种丝氨酸蛋白酶抑制剂, 而OVALY蛋白就是其一[18]。这些蛋白是鸡蛋结构的重要组成成分(蛋壳、蛋清、蛋黄、卵黄膜等), 其生理功能还需要深入研究[19]。有研究显示, 这些蛋白与哺乳动物的对应蛋白具有一定的同源性, 可能参与了细胞增殖、卵泡血管形成、禽类胚胎体外发育、蛋壳生物矿化等[20]。在鸡受精卵孵化过程中, OVALY含量变化显著, 推测可能是胚胎的胃肠道蛋白酶或蛋黄蛋白酶的抑制剂[21]。鸽的OVALY蛋白是否具有丝氨酸蛋白酶抑制剂生物活性尚未知晓, 作为鸽卵中含量最高的蛋白, 可能还具有不为人知的潜在功能, 需要进行大量深入的研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|