{kind=link}
青藏高原裸大麦种质资源形态多样性分析
[徐肖 , 栾海业, 张英虎, 卢健, 乔海龙, 臧慧, 杨红燕, 沈会权
, 栾海业, 张英虎, 卢健, 乔海龙, 臧慧, 杨红燕, 沈会权* ]
, 栾海业, 张英虎, 卢健, 乔海龙, 臧慧, 杨红燕, 沈会权]
|
|


作者简介:徐肖(1992—),男,江苏姜堰人,硕士,研究实习员,主要从事麦类遗传育种研究。E-mail: xuxiao0913@foxmail.com
为了解青藏高原裸大麦区种质资源的遗传多样性,提高该地区裸大麦种质资源利用效率,对86份青藏裸大麦种质资源的形态多样性进行鉴定。结果表明:该地区裸大麦种质资源遗传多样性丰富,主穗长的形态多样性指数最高,为2.05;株型的遗传多样性指数最低,为0.47。总体表现为数量性状形态多样性指数大于质量性状。基于形态性状的聚类分析把86份材料聚为4类,第Ⅰ类是二棱种质,为高秆大穗大粒型,第Ⅱ类种质为矮秆多粒长穗型,第Ⅲ类种质为短穗型,第Ⅳ类种质为高秆多粒型。采用主成分分析进行种质评价,提取前7个主成分即可概括全部性状的大部分生物学信息,筛选出适合江苏地区生长的优良种质资源,为裸大麦种质资源的收集和利用提供理论依据。
To study the genetic diversity of hulless barley accessions from Qinghai-Tibetan Plateau and improve their utilization efficiency, the morphological diversity of 86 hulless barley accessions from Qinghai-Tibetan Plateau was identified in this study. The results showed that genetic diversity of hulless barley accessions was ample, morphological diversity index of main panicle length was the highest with the value of 2.05. Plant type had the lowest genetic diversity index with the value of 0.47. In general, genetic morphological index of quantitative traits were greater than those of the quality traits. The tested materials were divided into four groups by cluster analysis. Barley accessions in group Ⅰ were two-rowed, high plant height, big spike and big grain. Barley accessions in group Ⅱwere short plant height, multi grain and long spike. Barley accessions in group Ⅲ were short panicle. Barley accessions in group Ⅳ were high plant height and multi grain. Based on the principal components, comprehensive evaluation of various materials was carried out, seven principal components were extracted to summarize most of the biological information of all traits. Excellent germplasm resources suitable for growth in Jiangsu Province were screened out. This study provided a theoretical basis for the collection and utilization of hulless barley accessions.
裸大麦, 又名青稞, 是海拔高至4 000 m地区唯一能成熟的作物, 具有适应性广、早熟、抗逆性强等优点, 是高寒、干旱和沿海滩涂地区主要的粮食作物[1]。青藏高原裸大麦区种植面积达35.8× 104 hm2左右, 总量约99.4× 104 t, 至今藏族同胞仍以裸大麦为主食[2]。美国科学家于20世纪80年代研究发现, 大麦尤其是裸大麦β -葡聚糖具有降血脂, 降胆固醇, 预防糖尿病、心脑血管疾病, 排毒养颜, 提升免疫力等作用, 在调节血糖、抵御感染、抗肿瘤等方面也具有功效[3]。裸大麦β -葡聚糖作为非特异性的免疫调节剂, 可激活免疫细胞, 还能促进细胞生成, 激活补体系统, 增强抗体产生, 调节免疫系统。随着人们对裸大麦膳食纤维有益人体健康认识的不断增加, 高纤维食品越来越受到消费者的青睐。目前, 我国对裸大麦膳食纤维的研发与日本、欧美等发达国家还存在一定的差距, 随着我国人口老龄化的不断加剧, 糖尿病、心脑血管等疾病的预防越来越受到人们的重视。因此, 裸大麦膳食纤维的开发具有重要的现实意义和广阔的市场前景[3]。
裸大麦起源于东方栽培大麦, 在距今3 500~4 500年前通过巴基斯坦北部、印度和尼泊尔进入西藏南部[4]。基于裸大麦的重要性及其丰富的种质资源, 西藏自治区农牧学院与深圳华大基因科技服务有限公司已共同绘制完成裸大麦基因组图谱。用于基因组图谱构建的裸大麦品种是西藏古老的地方品种“ 拉萨钩芒” , 基因组大小约为4.5 Gbp[5]。青藏高原地区裸大麦种质资源类型和遗传多样性较丰富, 但关于该地区裸大麦种质资源的研究却报道不多, 胥婷婷[6]对402份青稞种质进行了形态和分子遗传多样性研究。本研究以86份青藏高原裸大麦种质资源为材料, 利用形态性状对其遗传多样性进行初步分析, 将这些裸大麦种质进行综合评价, 以便快速筛选出适合江苏沿海地区生长的综合性状优异的裸大麦种质, 为裸大麦种质资源的收集和利用提供理论依据。
以江苏沿海地区农业科学研究所收集的86份来自青藏高原地区的裸大麦品种(系)为参试材料。材料名称见表1。
| 表1 青藏高原裸大麦种质资源详情 Table 1 Details of hulless barley accessions from Qinghai-Tibetan Plateau |
于2016年、2017年秋播将试验材料种植于江苏沿海地区农业科学研究所南洋实验农场, 试验采用随机区组设计, 每个品种播种1行, 行长1.2 m, 行距25 cm, 株距3 cm, 每行点播40粒, 3次重复。因2017年种植材料在次年收获期出现大面积倒伏, 穗发芽严重, 导致考种数据不可用。
田间调查叶耳颜色、叶片姿势、株型、穗姿、棱型、小穗密度。收获时每品种取10株单株室内考种, 考种项目包括株高、主穗长、籽粒颜色、籽粒形状、每穗粒数、千粒重、结实率。调查和考种的主要形态性状和赋值标准如下。
株高(cm):10株株高平均。
主穗长(cm):10株主穗长平均。
每穗粒数:10株单株粒数平均。
千粒重(g):1 000粒籽粒质量, 重复2次取平均值。
叶耳颜色:1, 白; 2, 绿; 3, 红; 4, 紫。
叶片姿势:1, 直立; 2, 平展; 3, 下垂。
株型:1, 紧凑; 2, 半紧凑; 3, 松散。
穗姿:1, 直立; 2, 水平; 3, 下垂。
棱型:1, 二棱; 2, 中间型; 3, 六棱。
小穗密度:1, 稀; 2, 密; 3, 极密。
籽粒颜色:1, 黄; 2, 蓝; 3, 紫; 4, 褐; 5, 黑
籽粒形状:1, 长圆形; 2, 卵圆形; 3, 椭圆形; 4, 圆形。
结实率(%)=实粒数/(实粒数+空粒数)× 100。
遗传多样性指数[7](H’ , Shannon-Wiener diversity index):通过计算参试材料的平均数(μ )和标准差(σ ), 将考查的目标性状均划分为10级, 从第1级[xi< (μ -2σ )]到第10级[xi> (μ +2σ )], 每间隔0.5σ 为1级, 计算每一级相对频率pi, 从而得到多样性指数。计算公式为:
用SPSS 21.0统计软件进行聚类和主成分分析, 在聚类过程中, 种质间距离为欧氏距离, 聚类方法采用离差平方和法。
由表2可知, 参试材料主穗长的遗传多样性指数最高(2.05), 株型的遗传多样性指数最低(0.47), 变幅为0.47~2.05。说明青藏高原裸大麦种质资源的遗传多样性较为丰富, 且株高与主穗长的遗传多样性较高, 而株型与叶耳颜色的遗传多样性较低, 数量性状的遗传多样性普遍高于质量性状。遗传多样性较低的质量性状中, 绝大多数(83.72%)叶耳颜色为白色, 少数为绿色或紫色; 绝大多数(82.56%)株型为半紧凑, 其余为松散。
| 表2 青藏高原裸大麦种质资源主要性状遗传多样性指数 Table 2 Genetic diversity of main characters of hulless barley accessions from Qinghai-Tibetan |
对86份材料5个主要数量性状的基本统计分析结果(表3)表明, 不同材料间差异很大, 表现出显著的形态多样性。除结实率外, 其余4个主要农艺性状的变异系数均较大。表明在这4个性状上供选择的育种材料很丰富。其中, 变异系数最大的性状是主穗长(33.08%), 变异系数最小的性状是株高(10.56%)。其余两性状分别是每穗粒数(25.11%)与千粒重(13.73%)。说明青藏高原裸大麦种质资源在主穗长与每穗粒数上差异较大, 在株高与千粒重上差异较小。
| 表3 青藏高原裸大麦种质资源数量性状形态多样性统计 Table 3 Statistical morphological diversity of quantitative traits of hulless barley accessions from Qinghai-Tibetan |
86份裸大麦种质13个主要农艺性状的相关分析结果见表4:叶耳颜色与叶片姿态呈极显著正相关, 与棱型呈显著负相关; 株型与小穗密度、结实率均呈显著负相关; 穗姿与千粒重呈显著正相关; 棱型与穗姿、主穗长呈显著负相关, 与每穗粒数呈极显著正相关; 小穗密度与籽粒形状呈极显著正相关, 与棱型、结实率均呈显著正相关; 籽粒颜色与千粒重呈显著正相关, 与主穗长呈显著负相关; 籽粒形状与千粒重、结实率均呈极显著正相关; 千粒重与结实率呈显著正相关; 主穗长与每穗粒数呈极显著正相关; 株高与籽粒颜色呈极显著正相关, 与结实率、每穗粒数呈显著负相关。
| 表4 青藏高原裸大麦种质资源主要农艺性状的相关性 Table 4 Correlative coefficient of agronomic traits in hulless barley accessions from Qinghai-Tibetan |
以13个主要农艺性状(5个数量性状、8个质量性状)为指标, 基于马氏距离的类平均法, 使用R语言对86份青藏高原裸大麦种质进行聚类分析, 结果如图1所示。86份裸大麦种质在马氏距离10处可聚为4类。
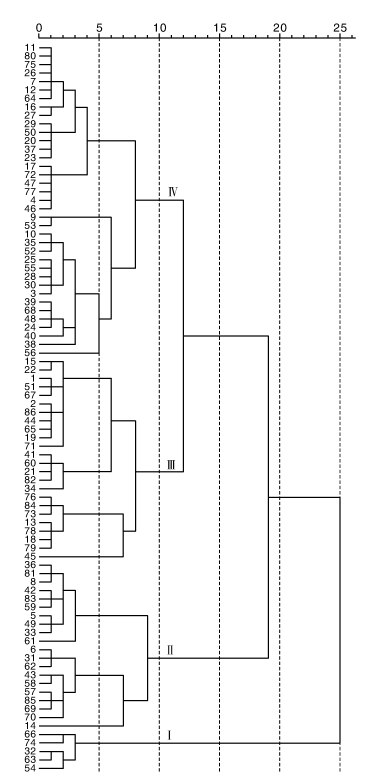 | 图1 青藏高原裸大麦种质聚类图 1~86为材料编号, 与表1同。Fig.1 UPGMA clustering dendrogram of hulless barley accessions from Qinghai-Tibetan Plateau ID of hulless barley accessions 1-86 were the same as that in table 1. |
第Ⅰ 类群包括5份材料, 该类群材料基本为二棱种质资源, 因此表现为植株较高, 平均穗长为4个类群中最长, 平均穗粒数为4个类群中最少, 穗粒重较高, 株型均呈半紧凑, 穗姿均呈直立型, 结实率较高, 属于大穗型品种, 综合表现一般。
第Ⅱ 类群包括20份材料, 该类群材料为六棱或中间型资源, 表现为株高适中、穗较长、穗粒数最多、穗粒重一般、结实率较高, 综合表现较好。
第Ⅲ 类群包括24份材料, 该类群材料表现为矮秆, 平均穗长为4个类群中最短, 穗粒重一般, 穗粒数较少, 结实率一般, 综合表现一般。
第Ⅳ 类包括37份材料, 该类群材料为六棱或中间型品种, 其特征为植株最高, 结实率最低, 千粒重最低, 穗粒数适中, 综合表现较差。
裸大麦种质各性状间存在不同程度的相关性, 因此为消除性状间相关性的影响, 本研究利用主成分分析进行种质评价。按照累计贡献率大于70%的原则, 提取前7个主成分即可概括全部性状的大部分生物学信息(表5)。
| 表5 青藏高原裸大麦主成分分析的特征值与贡献率 Table 5 Eigenvalue and rate of contribution of PCA in hulless barley accessions from Qinghai-Tibetan Plateau |
第一主成分的贡献率为16.547%, 籽粒形状、千粒重、小穗密度和结实率的特征向量值最大, 且均为正值。籽粒越饱满, 千粒重、小穗密度和结实率的增加均有利于高产, 故在实际应用中应保持较高状态。
第二主成分的贡献率为15.112%, 棱型的特征向量值最大, 棱型值为负。棱型增加, 每穗粒数增加, 主穗长减少, 不利于大穗大粒的形成, 因而在实际应用中应保持第二主成分适中。
第三主成分贡献率为12.594%, 籽粒颜色、每穗粒数和株高的特征向量值最大, 每穗粒数的值为正, 籽粒颜色和株高值为负。每穗粒数的增加有利于高产, 株高的降低减少倒伏性, 籽粒颜色加深, 穗长减少, 不利于高产, 故实际应用时应选择此成分适中偏高的。
第四主成分贡献率为9.170%, 株型的特征向量值较大, 且为正值。株型松散易造成倒伏, 不利于高产, 故实际应用时此主成分应略小为好。
第五主成分贡献率为8.924%, 叶耳颜色和叶片姿态的特征向量值较大, 且均为正值, 叶耳颜色的加深, 叶姿下垂, 不易促进光照的吸收, 不利于高产。故实际应用时此主成分也应略小为好。
第六主成分贡献率为7.271%, 穗长的特征向
量值最大, 且为正值, 穗长增长, 每穗粒数增加, 有利于产量的提高, 故在实际应用中应选择此成分适中偏高的品种。
第七主成分贡献率为6.886%, 穗姿的特征向量值最大, 且为正值, 穗姿下垂, 千粒重较重, 每穗粒数较多, 有利于高产, 故在实际应用中应选择此成分偏高的品种。
对裸大麦种质13个主要农艺性状数值进行标准化处理, 获得前7个主成分得分, 将这7个主成分得分进行归一化处理, 以贡献率计算各主成分权重系数, 分别为0.216 3、0.197 5、0.164 6、0.119 9、0.116 6、0.095 0和0.090 0, 利用前7个主成分的得分值与其对应的权重乘积之和作指标, 得到每份种质材料的综合得分(即M值), 对各种质材料进行综合评价, M值越高, 综合性状表现越好。裸大麦种质材料综合性状得分排前10位的分别为:小青稞、循化黑青稞、福9、藏830563、青6265WDM3344、9820、昆仑14、ICNBFB-6189EL.3gp、查袜、足捉春青稞。这些种质综合表现较好, 可加以利用。
种质资源的遗传多样性是育种工作的基础, 近些年为了提高育种水平, 育种家逐步注重提高育种基础材料的遗传多样性, 以扩大育成品种的遗传基础[8, 9]。遗传多样性指数已被广泛应用于各类作物的多样性评价中, 尤其是水稻、小麦和棉花等核心种质的多样性评价[10, 11, 12]。潘志芬等[13]和吴昆仑[14]对青藏高原青稞进行了遗传多样性分析, 结果表明, 青藏高原青稞遗传基础较广, 具有丰富的遗传多样性。原红军等[15]研究认为, 西藏青稞育成品种遗传多样性大于西藏野生大麦, 也大于西藏青稞地方品种。胥婷婷[6]以来源于青藏和云贵高原的402份青稞种质为材料, 利用42对SSR标记结合6个农艺性状分析了这些青稞种质资源的遗传多样性, 结果发现:402份青稞种质在农艺性状上表现出丰富的遗传多样性, 西藏青稞的多样性最为丰富, 其次是青海青稞; 主成分分析说明株高对千粒重和产量有较为重要的影响; 采用聚类取样法、随机取样法和最小距离逐步取样法, 从402份青稞种质中筛选出50份具有较高遗传多样性的青稞种质作为核心种质。本研究结果表明, 86份裸大麦主要农艺性状遗传多样性指数均较高, 数量性状的遗传多样性指数(1.62~2.05)高于质量性状(0.47~1.04), 说明本次引进的青藏裸大麦资源具有较高的丰富度和均匀度, 遗传多样性广泛, 且数量性状遗传变异更加丰富。8个质量性状中, 籽粒颜色多样性指数最大, 这与夏腾飞等[16]的研究结果一致。本研究超过一半(59.30%)的种质资源籽粒为褐色, 近三分之一(31.40%)种质资源籽粒为黄色, 紫色籽粒占4.65%, 籽粒颜色为蓝色和黑色的最少, 各占2.33%。各数量性状的变异系数中, 主穗长和每穗粒数相对较高, 分别为33.08%、25.11%, 说明这些性状具有较大的选择范围, 利用现有资源育成大穗大粒型、产量高的品种或品系成为可能。
刘亚楠等[17]对全球97份二棱大麦种质研究发现, 主穗长与主穗粒数呈极显著正相关, 穗长增长, 主穗粒数增多。夏腾飞等[16]研究认为, 青藏高原裸大麦每穗实粒数和千粒重呈极显著正相关, 株高与每穗实粒数呈显著负相关, 本研究结果与此一致。方彦杰等[18]研究47份美国大麦种质资源发现, 株高与穗粒数呈极显著正相关, 本研究表明, 株高与每穗粒数呈显著负相关。原因可能是青藏高原位于高海拔地区, 盐城位于沿海地区, 生态环境的差异导致种植于盐城的部分青藏高原裸大麦种质株高增加, 植株养分源有限, 每穗粒数减少, 这一现象基本符合“ 源库流理论” 。
尚毅等[19]对浙江省裸大麦地方品种聚类研究发现, 裸大麦和皮大麦分别聚为一大类, 与品种地理来源没有联系。贾巧君等[20]研究发现, 浙江赤霉病抗性不同的大麦地方品种聚类与品种来源地无关, 原因是浙江省面积小, 各区域有着相同或相似的生长环境或气候条件, 或各地方品种间的频繁交流, 导致这些地方品种的地域程度降低。Chen等[21]对上海的大麦地方品种研究发现, 二棱大麦和六棱大麦分别聚为一类, 表明二棱品种与六棱品种有遗传差异, 与本研究结果类似。
本研究通过对引进的86份青藏高原裸大麦种质资源农艺性状综合鉴定和评价, 以期筛选出生态适应性、高产、优质、抗逆性等方面适宜于江苏沿海地区生长的优良品系材料。优质的大麦种质, 应具有优良的表型性状和品质。表型性状虽受环境影响, 但性状的表型鉴定比较直观; 分子标记鉴定结果比较可靠, 但同一性状往往由多个基因控制, 基因又有主效和微效之分, 很难利用少数几个标记准确鉴定某一性状的表型。表型性状的变异在一定程度上仍能反映出各类群的亲缘关系和表型特征。
本研究运用的马氏距离类平均法聚类可消除性状间相关性对聚类结果的影响[22], 本文将86份裸大麦种质资源分为4类, 第Ⅰ 类群种质最少, 第Ⅳ 类的种质最多。第Ⅰ 类是二棱种质, 为高秆大穗大粒型, 第Ⅱ 类种质为矮秆多粒长穗型, 第Ⅲ 类种质为短穗型, 第Ⅳ 类种质为高秆多粒型。Ⅰ 、Ⅳ 类种质高秆性状需要改良, 第Ⅲ 类种质虽为短穗型, 穗粒数少, 产量低, 但从丰富基因来看, 可作为个别性状的改良亲本。
M值评价法可为育种家提供更直观、更方便的参考指标, 为今后裸大麦种质资源的收集和利用提供了理论依据。同时, 本研究利用M值评价法筛选出的10份裸大麦种质, 对江苏地区青稞育种工作中亲本的高效选择具有一定的指导意义。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|