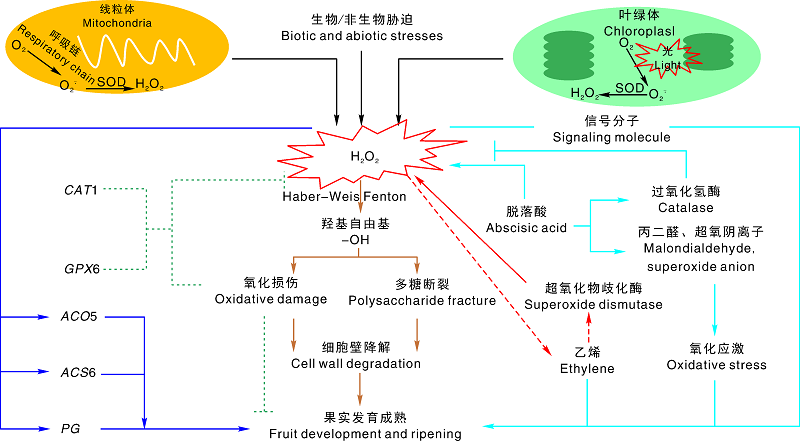
Advances in ROS promoting fruit development and ripening
WANG Zhenguang, YU Yihe, GUO Dalong*
College of Forestry, Henan University of Science and Technology, Henan Engineering Technology Research Center of Quality Regulation and Controlling of Horticultural Plants, Luoyang 471023, China
Abstract
ROS is produced along the fruit development, which often damages proteins, lipids, carbohydrates, and DNA, causing severe cell membrane damage and inducing programmed cell death. But moderate oxidative stress is conducive to fruit ripening. The physiological mechanism of ROS affecting the fruit ripening, the interaction mechanism of ROS and hormones regulating fruit ripening, the molecular mechanism of ROS directly regulating the fruit ripening, the cooperative interaction of ROS and calcium ions regulating postharvest fruit ripening were summarized, to provide a theoretical basis and reference for employing ROS to regulate fruit ripening directly or indirectly.
MITTLERR, VANDERAUWERAS, SUZUKIN, et al. ROS signaling: the new wave?[J]. Trends in Plant Science, 2011, 16(6): 300-309. [本文引用:2]
[2]
MILLERG, SUZUKIN, CIFTCI-YILMAZS, et al. Reactive oxygen species homeostasis and signalling during drought and salinity stresses[J]. Plant, Cell & Environment, 2010, 33(4): 453-467. [本文引用:1]
[3]
DASGUPTAN, BISWASP, KUMARR, et al. RETRACTED ARTICLE: Antioxidants and ROS scavenging ability in ten Darjeeling tea clones may serve as markers for selection of potentially adapted clones against abiotic stress[J]. Physiology and Molecular Biology of Plants, 2013, 19(3): 421-433. [本文引用:1]
[4]
PANDEY VP, SINGHS, JAISWALN, et al. Papaya fruit ripening: ROS metabolism, gene cloning, characterization and molecular docking of peroxidase[J]. Journal of Molecular Catalysis B: Enzymatic, 2013, 98: 98-105. [本文引用:2]
[5]
TSUKAGOSHIH, BUSCHW, BENFEY PN. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root[J]. Cell, 2010, 143(4): 606-616. [本文引用:1]
[6]
DENG XP, CHENG YJ, WU XB, et al. Exogenous hydrogen peroxide positively influences root growth and exogenous hydrogen peroxide positively influences root growth and metabolism in leaves of sweet potato seedlings[J]. Australian Journal of Crop Science, 2012, 6(11): 1572-1578. [本文引用:1]
[7]
LIAO WB, HUANG GB, YU JH, et al. Nitric oxide and hydrogen peroxide alleviate drought stress in marigold explants and promote its adventitious root development[J]. Plant Physiology and Biochemistry, 2012, 58: 6-15. [本文引用:1]
[8]
QUAN LJ, ZHANGB, SHI WW, et al. Hydrogen peroxide in plants: a versatile molecule of the reactive oxygen species network[J]. Journal of Integrative Plant Biology, 2008, 50(1): 2-18. [本文引用:2]
[9]
FASOLIM, DAL SANTOS, ZENONIS, et al. The grapevine expression atlas reveals a deep transcriptome shift driving the entire plant into a maturation program[J]. The Plant Cell, 2012, 24(9): 3489-3505. [本文引用:1]
[10]
GUO DL, XI FF, YU YH, et al. Comparative RNA-Seq profiling of berry development between table grape ‘Kyoho’ and its early-ripening mutant ‘Fengzao’[J]. BMC Genomics, 2016, 17(1): 1-17. [本文引用:2]
[11]
MØLLER IM, JENSEN PE, HANSSONA. Oxidative modifications to cellular components in plants[J]. Annual Review of Plant Biology, 2007, 58: 459-481. [本文引用:1]
[12]
SMIRNOFFN, ARNAUDD. Hydrogen peroxide metabolism and functions in plants[J]. New Phytologist, 2019, 221(3): 1197-1214. [本文引用:1]
[13]
BHATTACHARJEES. Reactive oxygen species and oxidative burst: roles in stress, senescence and signal transduction in plants[J]. Current Science, 2005, 89(7): 1113-1121. [本文引用:1]
[14]
SLESAKI, LIBIKM, KARPINSKAB, et al. The role of hydrogen peroxide in regulation of plant metabolism and cellular signalling in response to environmental stresses[J]. Acta Biochimica Polonica, 2007, 54(1): 39-50. [本文引用:1]
[15]
FOYER CH, LOPEZ-DELGADOH, DAT JF, et al. Hydrogen peroxide-and glutathione-associated mechanisms of acclimatory stress tolerance and signalling[J]. Physiologia Plantarum, 1997, 100(2): 241-254. [本文引用:1]
[16]
林毅雄, 林艺芬, 陈艺晖, 等. 过氧化氢对采后龙眼果实贮藏品质的影响[J]. 食品科学, 2016, 37(22): 244-248. LIN YX, LIN YF, CHEN YH, et al. Effects of hydrogen peroxide on quality of harvested longan fruits during storage[J]. Food Science, 2016, 37(22): 244-248. (in Chinese with English abstract)[本文引用:1]
[17]
MARINHO HS, REALC, CYRNEL, et al. Hydrogen peroxide sensing, signaling and regulation of transcription factors[J]. Redox Biology, 2014, 2: 535-562[本文引用:1]
[18]
HODGES DM, FORNEY CF. The effects of ethylene, depressed oxygen and elevated carbon dioxide on antioxidant profiles of senescing spinach leaves[J]. Journal of Experimental Botany, 2000, 51(344): 645-655. [本文引用:1]
[19]
HUANC, JIANGL, AN XJ, et al. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening[J]. Plant Physiology and Biochemistry, 2016, 104: 294-303. [本文引用:2]
[20]
PILATIS, BRAZZALED, GUELLAG, et al. The onset of grapevine berry ripening is characterized by ROS accumulation and lipoxygenase-mediated membrane peroxidation in the skin[J]. BMC Plant Biology, 2014, 14: 87. [本文引用:1]
[21]
LIN YF, LIN HT, ZHANGS, et al. The role of active oxygen metabolism in hydrogen peroxide-induced pericarp browning of harvested longan fruit[J]. Postharvest Biology and Technology, 2014, 96: 42-48. [本文引用:1]
[22]
LIN YF, LIN HT, LIN YX, et al. The roles of metabolism of membrane lipids and phenolics in hydrogen peroxide-induced pericarp browning of harvested longan fruit[J]. Postharvest Biology and Technology, 2016, 111: 53-61. [本文引用:1]
[23]
TODD JF, PALIYATHG, THOMPSON JE. Characteristics of a membrane-associated lipoxygenase in tomato fruit[J]. Plant Physiology, 1990, 94(3): 1225-1232. [本文引用:1]
[24]
JIMENEZA, CREISSENG, KULARB, et al. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening[J]. Planta, 2002, 214(5): 751-758. [本文引用:1]
[25]
KUMARV, IRFANM, GHOSHS, et al. Fruit ripening mutants reveal cell metabolism and redox state during ripening[J]. Protoplasma, 2016, 253(2): 581-594. [本文引用:4]
郭丽丽, 席飞飞, 余义和, 等. 核黄素处理促进‘巨峰’葡萄提早成熟的研究[J]. 园艺学报, 2017, 44(10): 1861-1870. GUO LL, XI FF, YU YH, et al. Studies of the riboflavin treatment for promoting the early ripening of ‘Kyoho’ Grape berry[J]. Acta Horticulturae Sinica, 2017, 44(10): 1861-1870. (in Chinese with English abstract)[本文引用:1]
[28]
INUPAKUTIKA MA, SENGUPTAS, DEVIREDDY AR, et al. The evolution of reactive oxygen species metabolism[J]. Journal of Experimental Botany, 2016, 67(21): 5933-5943. [本文引用:1]
[29]
DUAN XW, ZHANG HY, ZHANG DD, et al. Role of hydroxyl radical in modification of cell wall polysaccharides and aril breakdown during senescence of harvested longan fruit[J]. Food Chemistry, 2011, 128(1): 203-207. [本文引用:1]
[30]
CHENG GP, DUAN XW, SHIJ, et al. Effects of reactive oxygen species on cellular wall disassembly of banana fruit during ripening[J]. Food Chemistry, 2008, 109(2): 319-324. [本文引用:1]
[31]
ACHARDP, RENOU JP, BERTHOMÉR, et al. Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species[J]. Current Biology, 2008, 18(9): 656-660. [本文引用:1]
[32]
KUMARR, KHURANAA, SHARMA AK. Role of plant hormones and their interplay in development and ripening of fleshy fruits[J]. Journal of Experimental Botany, 2014, 65(16): 4561-4575. [本文引用:1]
[33]
FORLANIS, MASIEROS, MIZZOTTIC. Fruit ripening: the role of hormones, cell wall modifications, and their relationship with pathogens[J]. Journal of Experimental Botany, 2019, 70(11): 2993-3006. [本文引用:1]
[34]
ZHANG HB, LIA, ZHANG ZJ, et al. Ethylene response factor TERF1, regulated by ETHYLENE-INSENSITIVE3-like factors, functions in reactive oxygen species (ROS) scavenging in tobacco ( Nicotiana tabacum L. )[J]. Scientific Reports, 2016, 6: 29948. [本文引用:1]
[35]
MASIAA. Superoxide dismutase and catalase activities in apple fruit during ripening and post-harvest and with special reference to ethylene[J]. Physiologia Plantarum, 1998, 104(4): 668-672. [本文引用:1]
[36]
WANG LB, WANGL, ZHANGZ, et al. Genome-wide identification and comparative analysis of the superoxide dismutase gene family in pear and their functions during fruit ripening[J]. Postharvest Biology and Technology, 2018, 143: 68-77. [本文引用:1]
[37]
MONDALK, SHARMA NS, MALHOTRA SP, et al. Antioxidant systems in ripening tomato fruits[J]. Biologia Plantarum, 2004, 48(1): 49-53. [本文引用:1]
[38]
LARRIGAUDIÈREC, VILAPLANAR, SORIAY, et al. Oxidative behaviour of Blanquilla pears treated with 1-methylcyclopropene during cold storage[J]. Journal of the Science of Food and Agriculture, 2004, 84(14): 1871-1877. [本文引用:1]
[39]
WANG BG, WANG JH, FENG XY, et al. Effects of 1-MCP and exogenous ethylene on fruit ripening and antioxidants in stored mango[J]. Plant Growth Regulation, 2008, 57(2): 185-192. [本文引用:1]
[40]
HURR BM, HUBER DJ, VALLEJOS CE, et al. Ethylene-induced overproduction of reactive oxygen species is responsible for the development of watersoaking in immature cucumber fruit[J]. Journal of Plant Physiology, 2013, 170(1): 56-62. [本文引用:1]
[41]
LI TT, LI ZR, HU KD, et al. Hydrogen sulfide alleviates kiwifruit ripening and senescence by antagonizing effect of ethylene[J]. Hortscience, 2017, 52(11): 1556-1562. [本文引用:1]
[42]
陈昆松, 许文平. 脂氧合酶、茉莉酸和水杨酸对猕猴桃果实后熟软化进程中乙烯生物合成的调控[J]. 植物生理学报, 2000, 26(6): 507. CHEN KS, XU WP. Regulations of lipoxygenase, jasmonic acid and salicylic acid on ethylene biosynthesis in ripening kiwifruit[J]. Acta Phytophysiologica Sinica, 2000, 26(6): 507. (in Chinese)[本文引用:1]
[43]
YE NH, ZHU GH, LIU YG, et al. ABA controls H2O2 accumulation through the induction of OsCATB in rice leaves under water stress[J]. Plant & Cell Physiology, 2011, 52(4): 689-698. [本文引用:2]
[44]
GUAN LM, ZHAOJ, SCANDALIOS JG. Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response[J]. The Plant Journal: for Cell and Molecular Biology, 2000, 22(2): 87-95. [本文引用:1]
[45]
李红卫, 韩涛, 李丽萍, 等. ABA、GA3处理对冬枣采后果肉活性氧代谢的影响[J]. 园艺学报, 2005, 32(5): 793-797. LI HW, HANT, LI LP, et al. Effect of ABA and GA3 treatments on the metabolism of active oxygen species in cold stored ‘Brumal Jujube’ flesh[J]. Acta Horticulturae Sinica, 2005, 32(5): 793-797. (in Chinese with English abstract)[本文引用:1]
[46]
WANG JF, ZHANGL, CAO YY, et al. CsATAF1 positively regulates drought stress tolerance by an ABA-dependent pathway and by promoting ROS scavenging in cucumber[J]. Plant and Cell Physiology, 2018, 59(5): 930-945. [本文引用:1]
[47]
CHOUDHARYR, SAROHA AE, SWARNKAR PL. Effect of abscisic acid and hydrogen peroxide on antioxidant enzymes in Syzygium cumini plant[J]. Journal of Food Science and Technology, 2012, 49(5): 649-652. [本文引用:1]
[48]
ZHI HH, LIU QQ, DONGY, et al. Effect of calcium dissolved in slightly acidic electrolyzed water on antioxidant system, calcium distribution, and cell wall metabolism of peach in relation to fruit browning[J]. The Journal of Horticultural Science and Biotechnology, 2017, 92(6): 621-629. [本文引用:1]
[49]
KHALIQG, MUDA MOHAMED MT, GHAZALI HM, et al. Influence of gum Arabic coating enriched with calcium chloride on physiological, biochemical and quality responses of mango ( Mangifera indica L. ) fruit stored under low temperature stress[J]. Postharvest Biology and Technology, 2016, 111: 362-369. [本文引用:1]
[50]
KANG RY, YU ZF, LU ZX. Effect of coating and intermittent warming on enzymes, soluble pectin substances and ascorbic acid of Prunus persica(cv. Zhonghuashoutao) during refrigerated storage[J]. Food Research International, 2005, 38(3): 331-336. [本文引用:1]
[51]
BLOKHINAO, VIROLAINENE, FAGERSTEDT KV. Antioxidants, oxidative damage and oxygen deprivation stress: a review[J]. Annals of Botany, 2003, 91: 179-194. [本文引用:1]
[52]
段风琴. 壶瓶枣裂果的钙素营养生理及施钙效果研究[D]. 太谷: 山西农业大学, 2016. DUAN FQ. Influence of calcium on fruit cracking of jujube ‘Huping’ and its physiological mechanism[D]. Taigu: Shanxi Agricultural University, 2016. (in Chinese with English abstract)[本文引用:1]
[53]
KOU XH, GUO WL, GUO RZ, et al. Effects of chitosan, calcium chloride, and pullulan coating treatments on antioxidant activity in pear cv. “Huang guan” during storage[J]. Food and Bioprocess Technology, 2014, 7(3): 671-681. [本文引用:1]
[54]
KOU XH, WU MS, LIL, et al. Effects of CaCl2 dipping and pullulan coating on the development of brown spot on ‘Huangguan’ pears during cold storage[J]. Postharvest Biology and Technology, 2015, 99: 63-72. [本文引用:1]
[55]
韩絮舟, 吕静祎, 白琳, 等. 采后氯化钙处理对红树莓保鲜的影响[J]. 食品工业科技, 2020, 41(6): 233-238. HAN XZ, LYU JY, BAIL, et al. Effect of postharvest calcium chloride treatment on preservation of red raspberry[J]. Science and Technology of Food Industry, 2020, 41(6): 233-238. (in Chinese with English abstract)[本文引用:1]
[56]
赵妍, 杨超, 王若兰, 等. CaCl2处理对草莓采后品质及灰霉病害的影响[J]. 食品工业科技, 2013, 34(13): 313-316. ZHAOY, YANGC, WANG RL, et al. Influence of CaCl2 on quality and gray mold rot in postharvest strawberry fruit[J]. Science and Technology of Food Industry, 2013, 34(13): 313-316. (in Chinese with English abstract)[本文引用:1]
[57]
韩斯, 孟宪军, 汪艳群, 等. 氯化钙处理对速冻蓝莓冻藏期品质的影响[J]. 食品科学, 2014, 35(22): 310-314. HANS, MENG XJ, WANG YQ, et al. Effect of calcium chloride treatment on quality of quick frozen blueberry during frozen storage[J]. Food Science, 2014, 35(22): 310-314. (in Chinese with English abstract)[本文引用:1]
[58]
CHOUDHURY FK, RIVERO RM, BLUMWALDE, et al. Reactive oxygen species, abiotic stress and stress combination[J]. The Plant Journal, 2017, 90(5): 856-867. [本文引用:1]
[59]
HAMEEDA, GOHERM, IQBALN. Drought induced programmed cell death and associated changes in antioxidants, proteases, and lipid peroxidation in wheat leaves[J]. Biologia Plantarum, 2013, 57(2): 370-374. [本文引用:1]
[60]
PILATIS, PERAZZOLLIM, MALOSSINIA, et al. Genome-wide transcriptional analysis of grapevine berry ripening reveals a set of genes similarly modulated during three seasons and the occurrence of an oxidative burst at vèraison[J]. BMC Genomics, 2007, 8(1): 1-22. [本文引用:1]
[61]
APELK, HIRTH. REACTIVE OXYGEN SPECIES: metabolism, oxidative stress, and signal transduction[J]. Annual Review of Plant Biology, 2004, 55(1): 373-399. [本文引用:1]
[62]
ENDOH, OSEK, BAI JH, et al. Effect of hot water treatment on chilling injury incidence and antioxidative responses of mature green mume ( Prunus mume) fruit during low temperature storage[J]. Scientia Horticulturae, 2019, 246: 550-556. [本文引用:1]
[63]
MITTLERR. Oxidative stress, antioxidants and stress tolerance[J]. Trends in Plant Science, 2002, 7(9): 405-410. [本文引用:1]
[64]
GAOH, ZHANG ZK, CHAI HK, et al. Melatonin treatment delays postharvest senescence and regulates reactive oxygen species metabolism in peach fruit[J]. Postharvest Biology and Technology, 2016, 118: 103-110. [本文引用:2]
[65]
ZHENG XL, TIAN SP, MENG XH, et al. Physiological and biochemical responses in peach fruit to oxalic acid treatment during storage at room temperature[J]. Food Chemistry, 2007, 104(1): 156-162. [本文引用:1]
[66]
MARINOD, DUNANDC, PUPPOA, et al. A burst of plant NADPH oxidases[J]. Trends in Plant Science, 2012, 17(1): 9-15. [本文引用:1]
[67]
ZHUZ, CHEN YL, SHI GQ, et al. Selenium delays tomato fruit ripening by inhibiting ethylene biosynthesis and enhancing the antioxidant defense system[J]. Food Chemistry, 2017, 219: 179-184. [本文引用:1]
[68]
MUÑOZ-VARGAS MA, GONZÁLEZ-GORDOS, CAÑASA, et al. Endogenous hydrogen sulfide (H2S) is up-regulated during sweet pepper ( Capsicum annuum L. ) fruit ripening. In vitro analysis shows that NADP-dependent isocitrate dehydrogenase (ICDH) activity is inhibited by H2S and NO[J]. Nitric Oxide, 2018, 81: 36-45. [本文引用:1]
[69]
CHAKIM, ÁLVAREZ DE MORALESP, RUIZC, et al. Ripening of pepper ( Capsicum annuum) fruit is characterized by an enhancement of protein tyrosine nitration[J]. Annals of Botany, 2015, 116(4): 637-647. [本文引用:1]
[70]
LIT, ZHANGJ, GAO XY, et al. The molecular mechanism for the ethylene regulation of postharvest button mushrooms maturation and senescence[J]. Postharvest Biology and Technology, 2019, 156: 110930. [本文引用:1]
[71]
ZERMIANIM, ZONINE, NONISA, et al. Ethylene negatively regulates transcript abundance of ROP-GAP rheostat-encoding genes and affects apoplastic reactive oxygen species homeostasis in epicarps of cold stored apple fruits[J]. Journal of Experimental Botany, 2015, 66(22): 7255-7270. [本文引用:1]
[72]
MITTLERR, VANDERAUWERAS, GOLLERYM, et al. Reactive oxygen gene network of plants[J]. Trends in Plant Science, 2004, 9(10): 490-498. [本文引用:1]
[73]
ZHANGY, GONGY, CHENL, et al. Hypotaurine delays senescence of peach fruit by regulating reactive oxygen species metabolism[J]. Scientia Horticulturae, 2019, 253: 295-302. [本文引用:1]
[74]
GUO DL, ZHANG GH. A new early-ripening grape cultivar-‘fengzao’[J]. Acta Horticulturae, 2015(1082): 153-156. [本文引用:1]
[75]
CHIANG CM, CHIEN HL, CHEN L FO, et al. Overexpression of the genes coding ascorbate peroxidase from Brassica campestris enhances heat tolerance in transgenic Arabidopsis thaliana[J]. Biologia Plantarum, 2015, 59(2): 305-315. [本文引用:1]
林毅雄, 林艺芬, 陈艺晖, 等. 过氧化氢对采后龙眼果实贮藏品质的影响[J]. 食品科学, 2016, 37(22): 244-248. LIN YX, LIN YF, CHEN YH, et al. Effects of hydrogen peroxide on quality of harvested longan fruits during storage[J]. Food Science, 2016, 37(22): 244-248. (in Chinese with English abstract)
Excessive generation of reactive oxygen species (ROS) has been identified as a major cause of quality deterioration of harvested fruit and vegetables. The effect of hydrogen peroxide (H2O2), the most stable of the reactive oxygen, on the quality of harvested longan (Dimocarpus longan Lour.) fruit during storage were investigated. The results showed that in comparison to the control longans, the H2O2-treated longans had lower contents of pericarp pigments including chlorophyll, carotenoid and flavonoid, lower contents of pulp nutrients such as total sugar, sucrose and vitamin C, higher pericarp browning index and aril breakdown index, and lower percentage of commercially acceptable fruits. From the above findings, it could be concluded that H2O2 may play a key role in causing the quality deterioration of harvested longan fruit by accelerating the deterioration of pericarp color, stimulating the degradation of pulp nutrients, thereby reducing the marketable value of harvested longan fruit.
Institute of Postharvest Technology of Agricultural Products, College of Food Science, Fujian Agriculture and Forestry University, Fuzhou 350002, China
李红卫, 韩涛, 李丽萍, 等. ABA、GA3处理对冬枣采后果肉活性氧代谢的影响[J]. 园艺学报, 2005, 32(5): 793-797. LI HW, HANT, LI LP, et al. Effect of ABA and GA3 treatments on the metabolism of active oxygen species in cold stored ‘Brumal Jujube’ flesh[J]. Acta Horticulturae Sinica, 2005, 32(5): 793-797. (in Chinese with English abstract)
The changes of active oxygen species in ‘Brumal Jujube’flesh treated with 100 mg·L - 1 ABA or 100μg·L - 1 GA 3 during cold storage 〔(0 ±2) ℃, 60 days〕, including the p roduction rate of superoxide anion, contents of malondialdehyde (MDA ) or H 2 O 2 and activities of catalase (CAT) , peroxidase ( POD ) , polyphenoloxidase ( PPO ) and ascorbic acid oxidase (AAO ) , were investigated. The results showed that ABA increased the p roduction rate of superoxide anion, promoted the accumulations of MDA and H 2 O 2 in ‘Brumal Jujube’during storage, the activity peaks of CAT, PPO and POD were advanced, and AAO activity kep t increasing, the content of total phenols decreased sharp ly, all of the parameters above were higher than those in control. They suggested that ABA treatment accelerated the senescence of‘Brumal Jujube’fruit. The effects of ABA on ‘Brumal Jujube’fruit were opposite to GA 3 .
( 1 Department of Food Science, Beijing Agricultural College, Beijing 102206, China; 2 College of Food Science and Nutrional Engineering, China Agricultural University, Beijing 100083, China)
韩斯, 孟宪军, 汪艳群, 等. 氯化钙处理对速冻蓝莓冻藏期品质的影响[J]. 食品科学, 2014, 35(22): 310-314. HANS, MENG XJ, WANG YQ, et al. Effect of calcium chloride treatment on quality of quick frozen blueberry during frozen storage[J]. Food Science, 2014, 35(22): 310-314. (in Chinese with English abstract)
Blueberry fruits were treated with varying concentrations of calcium chloride before storage at -18 ℃ for 6 months, and changes in blueberry quality were investigated during the storage period. The results indicated that calcium chloride resulted in a decrease in weight, firmness, total soluble solid (TSS), titratable acid, vitamin C, polyphenol and anthocyanin in blueberry fruits during the frozen storage. Calcium chloride treatment also retarded the decrease of fruit firmness, TSS, polyphenol, anthocyanin and vitamin C. In addition, the soluble polyphenol oxidase (PPO) activity and soluble peroxidase (POD) activities were decreased. Calcium chloride treatment significantly inhibited PPO activity in the later period of frozen storage, but had no significant effect on POD activity. By comprehensive analysis, 1.5% calcium chloride treatment on quick frozen blueberry fruits presents the optimum protection effects during frozen storage.
College of Food Science, Shenyang Agricultural University, Shenyang 110866, China
{kind=link}
 , 余义和, 郭大龙
, 余义和, 郭大龙