


{kind=link}
{kind=link}
光周期调控植物开花分子机制以及CCT基因家族研究进展
[袁玺垒 , 王振山, 贾小平
, 王振山, 贾小平* , 桑璐曼, 李剑峰, 张博]
, 王振山, 贾小平, 桑璐曼, 李剑峰, 张博]
|
|


作者简介:袁玺垒(1993—),男,河南项城人,硕士研究生,研究方向为植物分子细胞生物学。E-mail : yuan_xilei@163.com
植物从营养生长到生殖生长的成花转变是其适应外界环境确保自身繁衍的关键环节,主要受内部调控和外部环境因素的影响,其中光周期是影响植物开花的一个重要因素,目前对其调控开花的分子机制研究也较为深入。本文结合近几年最新研究成果,总结了拟南芥、水稻、高粱和谷子4种植物光周期调控开花的分子机制,在此基础上还介绍了在植物光周期途径中扮演重要角色的一类含有CCT结构域的基因家族,对其不同成员在各个作物中的功能进行了概述,以期为进一步研究CCT基因家族的功能和开展光周期调控开花相关性状的分子育种提供理论基础。
Flowering transformation from vegetative growth to reproductive growth is the key link for plants to adapt to the external environments and ensure their reproduction, which is mainly influenced by internal regulation and external environmental factors. Photoperiod is an important factor that affects plant flowering, and the researches on molecular mechanism of photoperiod-regulation flowering were relatively in-depth. In this paper, the molecular mechanism underlying photoperiod-regulation flowering in Arabidopsis, rice, sorghum and foxtail millet was summarized combined with the latest research achievements in recent years. Furthermore, a gene family containing CCT motif that played a key role in photoperiod-regulation flowering pathway was introduced, and the functions of different members of this family in each crop were generalized, to provide theoretical reference for further study on the function of CCT motif-containing gene family and molecular breeding of traits related to photoperiod-regulated flowering.
植物从营养生长到生殖生长的成花转变是适应外界环境确保自身繁衍的关键环节, 主要受内部调控和外部环境的共同作用[1]。植物开花调控主要涉及光周期途径、春化途径、自主途径和赤霉素途径[2, 3]。随着相关研究进一步深入, 年龄途径、细胞分裂素途径、油菜素内酯途径、温敏途径、糖代谢等新的开花调控途径也相继被发现[4, 5, 6, 7, 8, 9, 10, 11]。光周期是影响植物生长发育的一个非常关键的环境因素, 有关光周期调控植物开花的分子机理在拟南芥、水稻中有大量报道, 特别是作为禾本科模式作物的水稻, 其光周期研究成果对玉米、高粱、谷子等农作物研究具有重要借鉴意义。本文以拟南芥、水稻、高粱、谷子4种植物为代表, 概述了光周期调控开花途径最新研究进展以及CCT结构域基因家族在植物开花调控中所起的关键作用。
拟南芥作为经典的长日照植物, 在长日照状态下能提前开花, 短日照抑制开花, 光周期调控开花途径已经十分清晰, 也比较完善, 为其他作物相关研究提供了重要的参考[12]。拟南芥的开花分子调控网络有3个非常重要的开花调控因子, 分别是FLOWERING LOCUS T(FT)、LEAFY(LFY)和SUPPRESSOR OF OVEREXPRESSION OF CO 1(SOC1)。FT是一种开花因子, 受到上游基因CONSTANS(CO)的直接调控, 同时FT还受到春化途径和自主途径中FLC基因的调控, 主要在叶片中表达后, 通过转运至植株茎的上部分, 从而激活花的分生组织特异性基因的表达, 促使植株抽穗开花[13, 14]。在光周期途径中通过光敏色素对光信号的感知, 感受光照和黑暗的长短以及光的强弱程度, 产生相应的反应, 形成昼夜节律, 如果昼夜长度发生改变, 就会破坏已有的平衡, 导致某些开花的基因表达受到促进或抑制, 从而导致开花推迟或者提前[15]。例如, 植株叶片中的光敏色素A(phyA)、光敏色素B(phyB)、隐花色素2(CRY2), 它们直接或间接调节CO的表达, 然后传递给生物钟蛋白, 节律性地激活FT的表达, 从而诱导开花[16, 17]。在长日照条件下, GI和CO能够促进FT的表达, 使拟南芥提前开花, 其中CO基因是一个关键光周期基因, 受到多个因子的共同调控, 可以整合光信号和生物钟信号, 调控各种蛋白之间的相互作用, 从而起到抑制或促进作用[18, 19]。长日照条件下, 拟南芥的CO突变体开花推迟, 而CO功能正常的植株能促进开花; 短日照条件下, 拟南芥不能开花, 而CO的超表达可以使拟南芥在任何光周期条件下都提前开花, 说明大量CO蛋白的积累是拟南芥开花所必需的, CO可以正向调节拟南芥的开花时间[20, 21]。
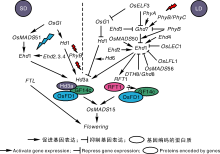
水稻作为禾本科的模式作物, 与拟南芥刚好相反, 是兼性短日照植物, 使得其光周期调控开花的途径与拟南芥既有相同之处, 又在一定方面存在差异。通过QTL定位方法已经在水稻中成功克隆了大量光周期途径中调节控制开花的关键基因, 如Heading date-3a (Hd3a)、Heading date 1 (Hd1)、Early Heading date(Ehd1)、Grains.Height.Date-7(Ghd7)等[22, 23, 24, 25]。水稻中开花相关基因Hd1与拟南芥CO是同源基因, 在表达规律上两者具有相同的昼夜节律, 而且中午有一个峰值[23]; 水稻Hd3a基因是拟南芥FT的同源基因, 是一种可以移动的开花信号, 被称为成花素, Hd3a起初在叶片中表达, 编码蛋白从叶片转移到顶端分生组织后, 诱导相关基因来促进花器官的发育, 由此行使与FT相同的功能[26]。短日照条件下, 水稻叶片感知短日照信号后, 主要通过Hd1途径和Ehd1途径调控Hd1和Ehd1的表达, 从而促进成花素Hd3a的表达, 作用于生殖发育, 加速植物开花。长日照条件下, Hd3a基因表达受到抑制, 导致开花推迟, 也有两条途径。长日照条件下, 在OsGI超表达突变体中Hd1白天的表达量增加, 而白天生成的光敏色素信号会和Hd1蛋白质相互作用使Hd1抑制Hd3a的表达, 从而推迟开花[27]。Xue等[25]研究发现, 长日照诱导Ghd7的高表达, 导致Ehd1途径的表达受到抑制, 从而Hd3a基因表达减少, 使植物开花被推迟; 短日照下Ghd7在水稻中几乎不表达, 于是减弱了对Ehd1的抑制, Ehd1诱导Hd3a表达, 造成成花素大量积累, 从而缩短植株开花时间(图1)。
Song等[28]鉴定出一个OsMFT1(MOTHER OF FT AND TFL1)基因, 是一种抑制开花的调控因子, 可同时调控水稻的抽穗和穗型, 过表达OsMFT1显著抑制了Ehd1表达, 同时Ghd7上调OsMFT1的表达, 推迟开花, 显著增加了每穗颖花数, 并大大增加了分枝数。
一些光周期途径的关键基因同时也受温度调节, 宋远丽等[29]研究了水稻光周期开花相关的几个关键基因在不同光温处理下的表达模式, 发现低温(23 ℃)条件下Ehd1、Hd3a和RFT1表达受到抑制而降低, 导致水稻开花受到影响, 推迟开花; 而在长日照条件下, 低温(23 ℃)上调了Ghd7的表达, Ghd7蛋白大量积累, 成花素减少, 从而抽穗开花延迟, 这表明长日照和低温这两个因素都促进Ghd7的表达, 延迟水稻开花。
高粱作为一种典型的短日照C4禾本科作物, 其成花的转变也受到包括光周期、温度、赤霉素等多种因素共同影响。其中光周期途径调控的开花时间也是由一系列感光信号基因和成花素基因参与的光信号级联途径所控制, 并与昼夜节律共同作用[30]。目前, 高粱光周期调控基因已经有一部分被克隆, 其受光周期影响开花时间的分子调控机制也逐渐清晰。长日照条件下, 对光周期敏感的高粱品种的开花时间延迟, 主要受到由SbPRR37(Ma1)和SbGhd7(Ma6)的双重正向调控作用。在高粱中开花促进基因SbEhd1和SbCO诱导FT的同源基因SbCN8和SbCN12的表达来促进开花。而在长日照条件下, SbPRR37和SbGHD7通过抑制开花激活子SbEHD1基因的表达延迟开花[31, 32]; 同时, SbPRR37还抑制了高粱另一个开花激活子SbCO的活性, 从而下调其表达, 也导致开花延迟, 使高粱的营养生长持续时间长, 提高了生物量和氮利用效率[33]。研究表明, 当Ma3和Ma5基因同时以显性形式存在时, 高粱的感光度进一步增强; 高粱生物钟调控的核心基因TOC、CCA1和LHY的规律性表达, 激活SbPRR37和SbGHD7早间和晚间的表达, 导致开花时间更加延迟[31, 32]。而在短日照条件下, SbPRR37和SbGHD7的表达因光照的减少, 开花激活基因SbEHD1和SbCO的表达增加, 进而上调了成花素基因SbCN8和SbCN12的表达, 促进提早开花。另外, 高粱生物钟调控基因GI的表达, 能促进SbCO表达, 然后SbCO通过直接或者间接上调成花素基因SbCN8和SbCN12的表达, 从而促进开花(图2)[33]。
谷子是自花授粉二倍体(2n=2x=18)作物, 基因组较小(约515 Mb), 具有抗旱、耐瘠和高光效的优良品质, 是C4禾谷类模式作物的理想模型。谷子也是短日照作物, 对光周期较敏感。目前, 谷子全基因组序列已经测定[34, 35], 相比玉米、高粱等基因组较大的C4作物, 谷子更适合作为C4作物光周期调控开花途径研究的模型。然而谷子光周期相关研究报道极少。贾小平等[36]通过利用不同国家、不同生态区域的100份春谷和60份夏谷材料为研究对象, 系统评价春谷和夏谷10个性状的光温敏感性, 揭示不同光温条件对谷子的影响, 以及各性状对光温反应的敏感性, 筛选能够准确反映谷子光温敏感性的指标性状, 为开展谷子光温敏感性遗传学分析、光敏感性相关基因的定位奠定基础。在谷子光周期敏感相关性状QTL定位研究中, 在谷子第4号染色体定位到短日照条件与光周期敏感性相关的QTL位点, 在第3和第9号染色体定位到长日照条件下与光周期敏感相关的QTL位点[37, 38]。Liu等[39]通过对高粱、水稻、谷子进行比较基因组学研究, 发现在谷子4号染色体存在水稻Hd1基因的同源基因, 关联分析表明, 谷子Hd1中存在一个高频的从GT到AT的剪接位点变异, 该变异导致了谷子的生育期变长, Hd1基因在水稻、高粱、谷子中受平行驯化, 但是该研究没有阐明谷子Hd1基因相关的分子作用机理。
CCT结构域是在拟南芥中最早被报道的, 由拟南芥中CONSTANS(CO)蛋白、CO-LIKE蛋白和TIMING OF CABl (TOCl)蛋白C末端的43个同源保守氨基酸序列组成[40, 41]。根据CCT基因中结构域的数量和类别, 可划分为3个亚家族, 分别是CMF、COL和PRR亚家族, 其中COL亚家族则含有B-box锌指结构域和一个CCT结构域; PRR亚家族含有PRR结构域和一个CCT结构域; 而CMF亚家族只有CCT一个功能结构域[42]。Cockram等[43]通过对禾本科植物不同物种的CCT基因家族的CCT基因序列进行比对和系统发育分析, 结果表明, CMF亚家族是在单子叶植物/双子叶植物进化分支后, 从COL基因的共同祖先进化演变成的, 推测COL基因的持续进化导致B-box结构域消失。
许多CCT域基因都直接或者间接地调节植物开花时间, 参与生物钟调控, 是光信号途径中的重要组成部分[44, 45]。拟南芥的CCT域基因CO、TOC1、COL1、COL2、COL9等参与花期调控的遗传机制已基本清楚[46, 47, 48]。水稻首先被克隆出来的CCT域基因Hd1是拟南芥中CO的同源基因, 长日照处理可以推迟水稻抽穗开花, 而在短日照处理下促进水稻抽穗开花, 与拟南芥有着截然相反的功能[23]。另外, 不管是在长日照条件下还是短日照条件下, Hd1都节律性地在夜间达到一个表达峰值, 还可以通过影响每穗的小穗数(粒数)来控制水稻产量[49]。Wu等[50]克隆了一个CCT域基因DTH2, 属于COL家族, 在长日照条件下通过上调成花素基因Hd3a和RFT1的表达来促进水稻提前开花。薛为亚[51]克隆的一因多效基因Ghd7, 属于CMF家族, 其蛋白只有一个CCT结构域, 长日照下, Ghd7通过在Ehd1和Hd3a的上游起作用, 并抑制Ehd1和Hd3a的表达, 抑制水稻生殖发育, 影响着水稻的抽穗期、株高和产量。Liu等[52]通过对Ghd2进行研究发现, 该基因参与对叶片衰老和干旱的调节, 通过整合环境作为调节剂与不同蛋白质的相互作用, 将衰老过程信号转化为发育程序, 控制着谷粒数和株高。刘海洋[53]成功克隆一因多效基因Ghd7.1, 也称为OsPRR37或DTH7, 在叶片和幼穗中的相对表达比较高, 影响着水稻发育情况, 在长日照条件下Ghd7.1不影响Hd1的表达, 而对EHd1和Hd3a有显著的抑制作用。谭俊杰[54]研究发现, 1个定位在细胞核的CCT域基因OsCOL10, 在长日照和短日照条件下均抑制水稻开花, 还受到Ghd7的调控, 表明OsCOL10是水稻中的开花抑制子。
大麦、高粱和玉米中也有CCT域基因的研究报道。Campoli等[55]对大麦Hd1的同源基因HvCO1的研究中, 发现不管长短日照, HvCO1的过表达都上调了HvFT1的表达, 影响花序的发育和茎的伸长, 加速了开花时间, 使大麦提前开花, 然而却会使短日照下野生型大麦开花推迟。近期, Huang等[56]通过全基因组关联和靶向高分辨率连锁作图分析发现, 玉米的ZmCCT基因是水稻Ghd7的同源基因, 与光周期密切相关, 影响着昼夜表达模式。同时, Li等[57]的研究发现, ZmCCT也是qRfg1基因座内的因果基因, 对赤霉病和茎腐病有一定的抗性, 而且发现ZmCCT启动子中的一个CACTA转录元件可以抑制ZmCCT的表达, 从而降低光周期敏感性, 对促进开花有显著调控作用, 使得玉米在LD条件下得以传播种植。在高粱中, CO同源基因sbCO是COL亚家族一个成员, 含有B-box锌指结构域和一个CCT结构域, 在缺失sbGhd7和sbPRR37的背景下无论长日照还是短日照均能促进植株开花[58]。Murphy等[59]通过图位克隆得到Ma1基因, 也就是SbPRR37, 表达依赖于光, 受生物钟的调控, 在长日照条件下促进开花抑制子CO的表达, 进而抑制EHd1和FT的表达, 减少了EHd1和FT的积累而推迟开花。上述结果表明, CCT结构域基因虽然在不同物种中都保守地参与对开花时间的控制, 但是不同成员的具体功能与作用在物种间和物种内都发生了很大的分化[45]。
植物光周期调控开花的分子机制已经在拟南芥和水稻中获得了较大进展, 目前已经绘制出光周期开花途径的分子调控网络, 然而由于光周期调控机制极其复杂, 目前尚有许多参与光周期开花的基因没有被克隆, 因此还需要运用多种实验技术创制更多的突变材料, 克隆更多的参与光周期调控开花的新基因, 对目前的调控网络进行补充和扩展, 使人们对光周期开花途径有更深入、更细致的了解。此外, 植物开花受多种环境因素影响, 除了光周期, 环境温度也是影响植物开花的一个关键因子。已有研究表明, 许多参与光周期途径的基因也受到环境温度的影响, 光周期和环境温度存在明显的互作, 而对光温互作分子机制的研究较缺乏, 因此加强光周期和环境温度对植物开花的互作调控机制研究是未来的一个重要方向。一些光周期途径的关键基因, 如Ghd7、Ghd2等除了控制开花期外, 还对作物株高、穗部性状有显著影响, 因此在作物分子辅助选择育种中具有一定应用价值, 在进行光周期调控开花分子机理研究的同时, 也应该尝试将一些关键基因用于分子育种实践, 选育光周期钝感的适应性广、高产优质的作物新品种。最后需要说明的是, 目前水稻光周期途径研究较深入, 而作为C4作物的玉米、高粱、谷子等光周期调控途径研究较缓慢, 虽然在玉米、高粱中克隆了一些光周期途径的关键基因, 这些作物较大的基因组对进一步阐明C4作物光周期调控开花分子机制存在制约。谷子作为我国栽培历史悠久的传统作物, 遗传资源丰富, 存在较多的对光周期表现不同敏感性的自然变异材料, 加上较短的生育期、较少的水肥投入、相对小且已经完成测序的基因组, 使其成为一个理想的C4作物模型[60], 因此, 加强谷子光周期调控机理研究, 对C4作物光周期调控开花路径的阐明具有积极的促进作用。
(责任编辑 张 韵)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|