



{kind=link}
{kind=link}
{kind=link}
{kind=link}
不同类型烟草种质的烟碱含量变化与相关基因表达水平
[徐秀红 , 刘金亮, 李栋成, 刘仁祥
, 刘金亮, 李栋成, 刘仁祥* ]
, 刘金亮, 李栋成, 刘仁祥]
|
|


作者简介:徐秀红(1980—),女,安徽宣城人,博士,副教授,研究方向为种质创新与利用、作物分子改良。E-mail:5725861@qq.com
为研究不同类型烟草种质的烟碱含量变化规律与相关基因的表达情况,以烤烟K326、香料烟巴斯玛、白肋烟TN90、晾烟马里兰为材料,采用荧光定量PCR技术,测定了烟草不同生育时期上部叶、中部叶、下部叶的烟碱含量与烟碱代谢过程中相关基因 PMT、 QPT、 CYP82 E5 V2、 A622的表达水平。结果表明,随着生育期的推进,烟碱含量缓慢上升,打顶后急剧增加,除巴斯玛打顶后还继续上升外,K326、TN90和马里兰烟均表现为:打顶后烟碱含量急剧增加,后缓慢下降,再渐趋平稳。不同类型烟草种质的 PMT在不同生育期均有表达,苗期 PMT表达量较高,其变化趋势与烟碱含量的基本一致; QPT在烟草发育前期表达量较高,而后下降,至打顶前1周表达量出现一个峰值,打顶后表达量持续增加,与烟碱含量变化呈正相关; CYP82 E5 V2表达量在不同生育时期均维持在较高水平,呈波浪型,生长旺期时表达量上升,打顶前后均出现峰值,但巴斯玛成熟期时其表达量下降,打顶前 CYP82 E5 V2表达量与烟碱含量变化呈正相关,打顶后期至成熟期两者呈一定程度的负相关;K326和马里兰烟的 A622表达量呈波浪型上升,至成熟时仍维持较高水平,而TN90和巴斯玛在打顶前后1周表达量较高,后表达下降再上升,成熟时表达量较低,与烟碱含量变化无明显一致性。
In order to study the changes of nicotine content and expression of related genes in different types of tobacco germplasm, four tobacco types, including flue-cured tobacco K326, oriental tobacco Basma, burley tobacco TN90 and sun-cured tobacco Maryland were used as materials. Nicotine content in different leaves and the expression levels of nicotine metabolism related genes PMT, QPT, CYP82 E5 V2 and A622 were measured at different developmental stages. The results showed that with the passing of growth stage, the nicotine content increased slowly, and increased dramatically after topping. Except for Basma, the nicotine content continued to increase after topping, while in K326, TN90 and Maryland, the nicotine content increased sharply after topping, then decreased slowly, and gradually stabilized. PMT gene in different types of tobacco was expressed in different growth stages. Expression level of PMT was higher at the seedling stage, and its change trend was basically the same as that of the nicotine content. Expression level of QPT was higher in the early stage, then decreased, and reached a peak one week before topping. After topping, the expression level continued to increase, which was positively correlated with the change of nicotine content. Expression level of CYP82 E5 V2 remained at a high level in different growth stages, presenting a wave pattern, and increased in the bloom stage, and peaked before and after topping. However, expression level of CYP82 E5 V2 in Basma decreased when it was mature. Expression level of CYP82 E5 V2 was positively correlated with the change of nicotine content before the topping, and negatively correlated to a certain extent from the later topping stage to the mature stage. Expression level of A622 in K326 and Maryland rose in a wave-like manner, and remained at a high level at maturity, while in TN90 and Basma, expression level of A622 was higher one week before and after topping, then decreased, and increased again, the expression level was lower at maturity, and there was no obvious consistency with the change of nicotine content.
烟草属茄科草本植物, 是我国重要的经济作物之一, 在我国的种植面积广泛。烟叶质量直接影响其经济价值, 而烟碱含量又与烟叶质量密切相关, 是影响烟叶质量最重要的化学成分[1]。烟碱是烟草次生代谢的主要产物, 烟碱含量的高低直接影响烤后烟叶的吸食品质[2]。因此, 烟碱是评价烟草和卷烟质量的一项重要指标, 其合成代谢的途径目前已有较多研究[3]。参与烟碱合成代谢途径各个步骤的酶与编码酶的基因基本都已被克隆[4, 5, 6], 其中腐胺N-甲基转移酶(PMT)催化腐胺形成N-甲基腐胺, PMT控制着腐胺向烟碱合成方向的流动速率, 是烟碱合成代谢途径中的限速酶[7, 8]; 喹啉酸磷酸核糖转移酶(QPT)在吡啶类生物碱生物合成过程中控制烟酸的供应[7, 8]; 类异黄酮还原酶基因A622可能参与烟酸与甲基吡咯啉阳离子的缩合反应[6, 9]。此外, 还有一些转化基因如CYP82E5V2、CYP82E2、CYP82E10和CYP82E4[6, 10], 转运基因如MATE、JAT和NUP1[7], 调控因子如COI1和JAZ[7], 以及调控烟碱合成的转录因子, 如ERF转录因子家族[9, 11]和bHLH转录因子家族[12], 这些基因与烟碱含量密切相关。
不同烟草品种的遗传特性不同, 烟碱含量也存在差异。本文以烤烟K326、香料烟巴斯玛、白肋烟TN90、晾烟马里兰烟为材料, 选取成苗期、团棵期、旺长期、打顶前后、成熟期的上、中、下部叶片, 分析叶片烟碱含量变化, 以及烟碱合成代谢途径4个关键基因PMT、QPT、A622和CYP82E5V2的表达量, 为解释烟碱含量变化的分子机理提供理论基础。
烤烟K326、白肋烟TN90、香料烟巴斯玛(Basma)、晾烟马里兰烟(Maryland)由贵州大学烟草学院提供。试验试剂有RNeasy Plant Mini Kit(74904)试剂盒(QIANEN公司)、DEPC水(华大基因)、随机引物(Invitrogen)、RNA酶抑制剂(Invitrogen)、AMV(Invitrogen)、2× PCR MIX(QIAGEN)、TBE(华大基因)等。
本研究于2016年5— 9月在贵州大学磊庄基地(106° 55'E、26° 40'N)和贵州大学烟草学院(106° 67'E、26° 42'N)进行。试验材料种植于磊庄基地, 按随机区组设计, 每试验小区4行, 每行15株, 小区种植60株, 行株距1.10 m× 0.55 m, 按常规管理措施进行田间管理。
在不同发育时期(成苗期、团棵期、旺长期、打顶前后、成熟期)对烤烟的不同部位叶片(上部叶片、中部叶片、下部叶片)进行取样, 除苗期外, 其他时期的叶片分上、中、下3个部位取样。每3株样品混合为1个样本, 分别用于RNA提取。同时对采收的鲜烟叶进行杀青烘干制成烟叶样品, 用于烟碱含量测定。每个试验3个生物学重复。
1.3.1 烟碱含量
烟碱含量测定参照李春丽等[13]编著的《烟叶化学成分及分析》中的“ 紫外分光光度法” 进行。
1.3.2 RT-PCR分析
采用荧光定量PCR技术, 用RNeasy Mini Kit(74904)试剂盒提取总RNA, 按顺序在1.5 mL离心管中加入DEPC水6 μ L、RNA酶抑制剂(50 U· μ L-1)0.5 μ L、随机引物(50 pmol· μ L-1)2 μ L、RNA 2 μ L(体积10.5 μ L), 在65 ℃水浴处理5 min, 室温放置10 min, 高于8 000 × g离心5 s, 按顺序在1.5 mL离心管中加入RNA酶抑制剂(50 U· μ L-1)0.5 μ L、5× buffer 4 μ L、dNTP MIX(10 mmol· L-1)2 μ L、ddH2O 2 μ L、AMV(200 U· μ L-1)1 μ L(总体积20 μ L), 40 ℃水浴反应1 h, 90 ℃处理5~10 min, 水浴5 min, 8 000× g离心5 s, 把总RNA逆转录成cDNA。
在NCBI网站中(http://www.ncbi.nlm.nih.gov/)查询并获得所有相关基因的基因序列, 并依据已知序列设计引物(表1)。采用Rapid SYBR® Green试剂在ABI ViiA 7PCR仪上进行qRT-PCR分析, 反应体系(16 μ L):上、下游引物(50 pmol· μ L-1)各0.2 μ L、H2O 6.6 μ L、2× PCR MIX 8 μ L, cDNA 1 μ L。PCR反应条件:95 ℃ 2 min; 94 ℃ 10 s, 60 ℃ 10 s, 72 ℃ 40 s, 40个循环。每个样品做3次平行实验。反应Ct值的采集采用校正的阈值设定, 实时荧光定量PCR以actin作为内参基因, 以编号1的样本作为对照组, 采用2Δ Δ Ct法进行相对定量计算。
| 表1 用于cDNA扩增的PCR引物序列 Table 1 Sequences of the primers for cDNA amplification |
数据采用Excel 2010计算平均值、标准差, 采用SPSS 19.0软件、LSD法进行3因素方差分析和相关性分析, 用HEMI 2.0软件进行数据可视化分析。
在整个生育期, 不同类型烟草种质叶片中的烟碱含量为0.19%~3.73%(表2), 不同种质之间的烟碱含量存在差异。总体上, 烤烟K326的烟碱含量和香料烟巴斯玛的烟碱含量相对较低, TN90和马里兰烟两者的烟碱含量相对较高。在整个生育期, 4个类型的烟草种质烟碱含量变化规律也存在着差异。随着生育期推进, 烟碱含量缓慢上升, 打顶后急剧增加, 除巴斯玛打顶后还继续上升外, K326、TN90和马里兰烟均表现为:打顶后烟碱含量先增加, 后缓慢下降, 再渐趋平稳。
| 表2 四个类型烟草种质不同生育期的烟碱含量 Table 2 Nicotine content of four tobacco varieties leaves in different growth stages |
4个类型烟草上、中、下部叶片的烟碱含量随生育期的变化趋势基本一致。打顶前, 除TN90无明显规律外, K326、马里兰烟和巴斯玛不同部位叶片烟碱含量基本表现为下部叶> 中部叶> 上部叶; 打顶期, 除TN90外, 所有类型烟草各部位叶片的烟碱含量相当; 打顶后, 不同类型烟草各部位叶片的烟碱含量变化不一致, 马里兰烟各部位叶片的烟碱含量总体表现为上部叶=中部叶> 下部叶(表2)。生育期、叶片部位、烟草种质对烟碱含量的影响结果表明, 生育期、烟草种质对烟碱含量的影响达极显著水平(P< 0.01), 而叶片部位对烟碱含量影响不显著(P> 0.05)(表3)。
| 表3 生育期、叶片部位、烟草种质对烟碱含量、PMT、QPT、CYP82E5V2和A622基因相对表达量的影响 Table 3 Influence of growth stage, leaf location and tobacco germplasm on nicotine content and relative expression of PMT, QPT, CYP82E5V2 and A622 gene |
2.2.1 PMT基因
由图1可知, PMT基因在所有类型烟草的不同生育时期均有表达, 除K326外, TN90、马里兰烟、巴斯玛在苗期PMT基因表达量较高, 除巴斯玛打顶后表达量继续上升外, 其余类型均表现为打顶后表达量上升后再下降, 后趋于稳定。TN90、马里兰烟和巴斯玛的上、中、下部叶片PMT基因的表达量表现基本一致, 而K326上部叶片PMT基因的表达量在打顶前后与中、下部叶片不同(图1)。PMT基因相对表达量与烟碱含量呈显著正相关关系, 即该基因相对表达量越高, 烟碱含量越高(表4)。生育期、不同部位、烟草品种对PMT基因的相对表达量影响均不显著(P> 0.05), 表明PMT基因的相对表达量受环境、种质来源影响较小, 表达量相对稳定(表3)。
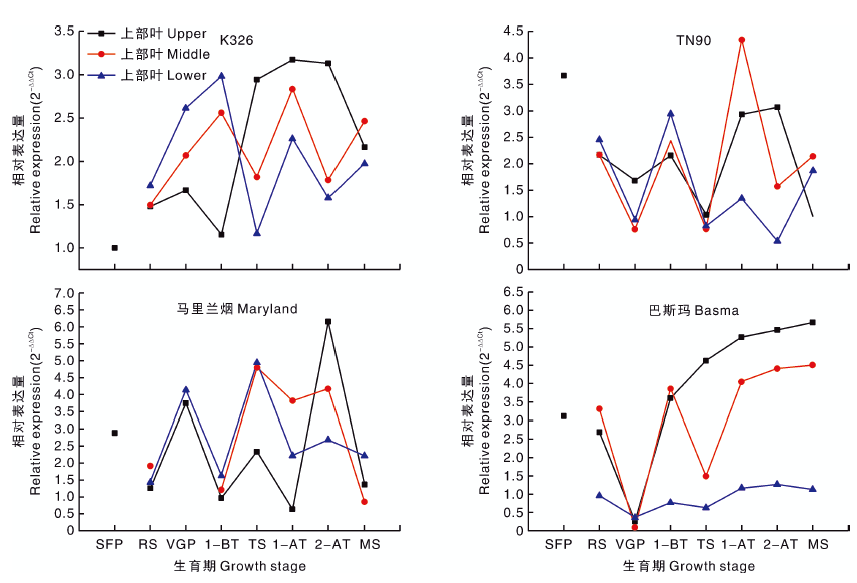 | 图1 四种类型烟草不同生育时期PMT基因的相对表达量 SFS, 成苗期; RS, 团棵期; VGS, 旺长期; 1-BT, 打顶前1周; TS, 打顶期; 1-AT, 打顶后1周; 2-AT, 打顶后2周; MS, 成熟期。下同。Fig.1 Relative expression level of PMT gene of four types of tobacco at different growth stages SFS, Seedling formation stage; RS, Rosette stage; VGS, Vigorous growing stage; 1-BT, 1 week before topping; TS, Topping stage; 1-AT, 1 week after topping; 2-AT, 2 weeks after topping; MS, Mature stage. The same as below. |
| 表4 烟碱合成代谢相关基因相对表达量与烟碱含量的相关性分析 Table 4 Correlation analysis of nicotine content and relative expression of the genes related to nicotinic anabolism |
2.2.2 QPT基因
4个类型烟草种质的QPT基因相对表达量变化较为一致, 在整个生育期呈波浪状起伏(图2)。在生育前期, 4个类型烟草种质的QPT基因表达量均相对较高, 除马里兰烟外, 其余类型均在打顶前1周出现峰值, 打顶后各类型烟草QPT表达量变化不一致。K326的QPT基因表达量在打顶后2周, 上部叶和下部叶出现峰值后下降, 而中部叶表达量持续上升。TN90上部叶和中部叶的QPT基因表达量在打顶后1周出现峰值, 之后略有下降再继续上升, 下部叶的QPT基因表达量在打顶后持续上升。马里兰烟下部叶和中部叶的QPT基因表达量在打顶后2周出现峰值, 然后下降, 上部叶则先下降后上升。巴斯玛在打顶后2周, 上部叶和下部叶的QPT基因表达量出现峰值后下降, 而中部叶的QPT基因表达量先下降后上升。QPT基因相对表达量受生育期、叶片部位影响, 且分别达显著(P< 0.05)、极显著水平(P< 0.01), 烟草种质对其影响不显著(表3)。QPT基因的表达量与烟碱含量无显著相关性(表4)。
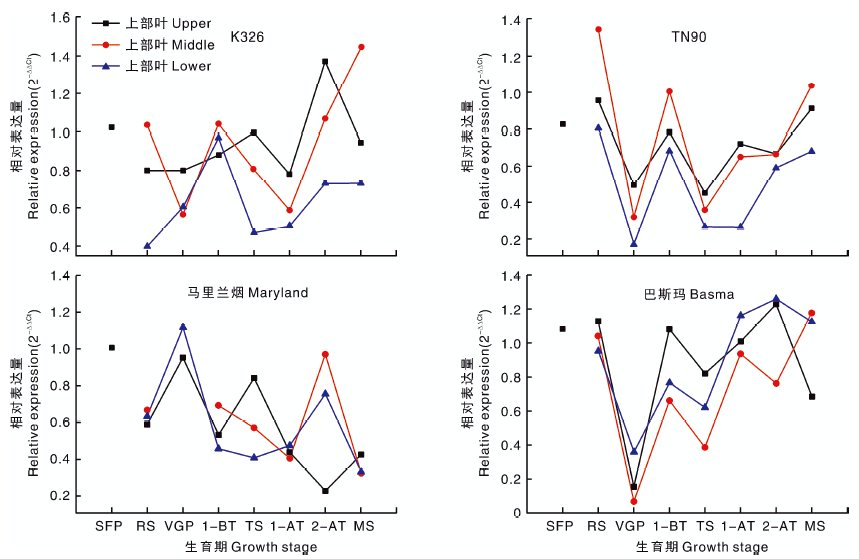 | 图2 四种类型烟草不同生育时期QPT基因的相对表达量Fig.2 Relative expression level of QPT gene of four types of tobacco at different growth stages |
2.2.3 CYP82E5V2基因
4个类型烟草不同部位的CYP82E5V2相对表达量随各自的生育期变化趋势基本一致, 仅K326上部叶片表达模式与其他不同(图3), K326的CYP82E5V2基因表达量则在打顶前1周和打顶后2周达到峰值, 说明打顶处理对CYP82E5V2的相对表达量产生了影响。生长发育时期、烟草种质对CYP82E5V2相对表达量的影响分别达极显著水平(P< 0.01)、显著水平(P< 0.05), 而不同部位对CYP82E5V2相对表达量无显著影响(P> 0.05)(表3)。其中, 马里兰烟中CYP82E5V2相对表达量与K326差异未达显著水平, 均高于TN90、巴斯玛。总体上, CYP82E5V2相对表达量在不同类型烟草种质的表现为K326> 马里兰烟> TN90> 巴斯玛, 且CYP82E5V2相对表达量与烟碱含量呈极显著正相关(表4)。
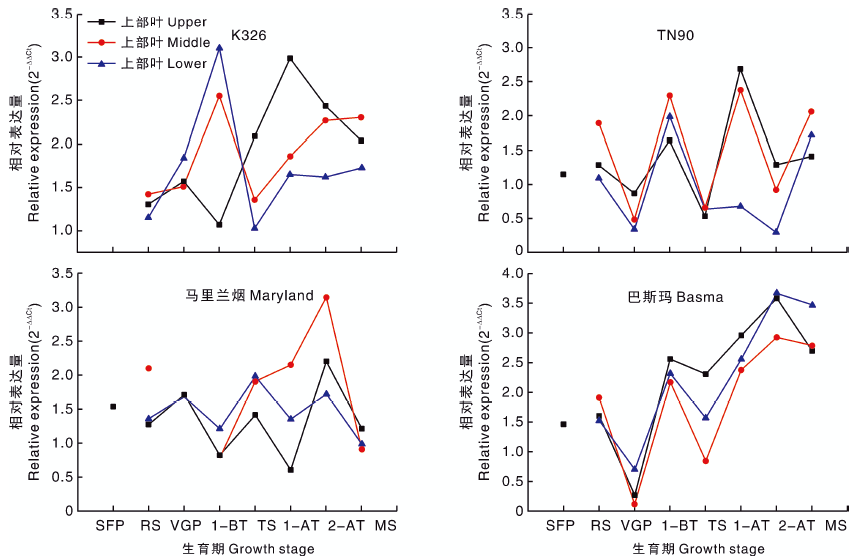 | 图3 四种类型烟草不同生育时期CYP82E5V2基因的相对表达量Fig.3 Relative expression level of CYP82E5V2 gene of four types of tobacco at different growth stages |
2.2.4 A622基因
K326、TN90和马里兰烟的A622不同生育期相对表达量表现为波浪状起伏; 而巴斯玛的A622表达量随生育期呈波浪状上升(图4), 在生长中前期(打顶前)相对较低, 打顶后至成熟期后逐渐升高, 特别是打顶后变化明显, 表明打顶处理对A622的相对表达量影响显著。生育期、烟草品种对A622表达量的影响达极显著水平(P< 0.01), 烟草种质对A622相对表达量的影响达显著水平(P< 0.05)(表3)。A622表达规律与烟碱含量变化规律基本一致, 其表达量与烟碱含量呈显著正相关的关系, 即该基因相对表达量越高, 烟碱含量越高(表4)。
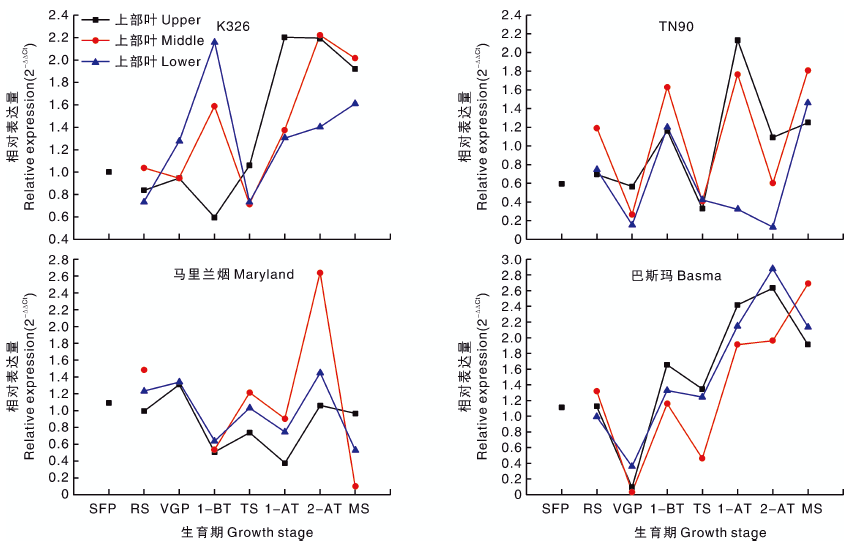 | 图4 四种类型烟草不同生育时期A622基因的相对表达量Fig.4 Relative expression level of A622 gene of four types of tobacco at different growth stages |
生物碱是烟草中最重要的一类化学成分, 生物碱的含量和比例决定烟叶感官品质与安全性。烟碱占烟草生物碱总含量的90%~95%, 占烟叶干重的0.6%~3.0%, 是评价烟草和卷烟质量的一项重要指标。烟碱含量的高低直接影响烤后烟叶的吸食品质, 过高会导致烟叶劲头偏大, 刺激性变强, 协调性变差。烟碱含量受很多因素影响, 除农艺栽培措施[14]、外源植物生长调节物质[15]、土壤和气候环境条件[16]等因素外, 烟草自身的遗传特性对其影响较大。本研究中的白肋烟TN90与晒晾烟马里兰烟种质来源接近, 两者烟碱含量也接近, 均高于烤烟K326和香料烟巴斯玛, 也说明了这一点。烟碱含量从成苗期到成熟期间的变化较大, K326、TN90在打顶前1周和打顶后1周烟碱含量处于较高水平, 巴期玛则持续增加, 在成熟期达到顶点, 且不同部位叶片烟碱变化趋势基本一致。
4个烟碱合成相关基因的相对表达量受烟草类型、发育时期、表达部位的影响不尽相同。PMT基因在整个生育期的相对表达量较为稳定, 受烟草类型、表达部位、发育时期的影响较小; QPT基因的相对表达量受发育时期、表达部位影响显著, CYP82E5V2相对表达量受生育期、烟草类型影响显著; A622的相对表达量受烟草种质类型、生育期和表达部位的影响显著。
烟碱的吡啶环部分由烟酸提供。在烟酸的生物合成途径中, QPT的作用是催化喹啉酸形成烟酰胺腺嘌呤二核苷酸, 因此QPT与烟碱的合成有较为直接的关联。本次研究中, 烟碱含量变化与QPT基因表达量的相关性不显著, 这表明QPT在烟碱生物合成过程中仅为烟酸的限速酶, 其表达量的高低不能直接决定烟碱的合成量, 其具体作用有待进一步研究。由于PMT是烟碱合成的关键酶[17, 18], 因此PMT基因表达量的高低直接决定了烟碱含量的高低。本研究结果也显示, 烟碱含量与PMT基因的表达量呈显著正相关关系。需要注意的是, 从烟草类型、生育期、表达部位看, PMT基因的表达始终处于较为稳定的水平, 受上述3种因素的影响不显著; 因此, PMT基因参与烟碱合成的途径与方式还需要深入研究。
不同类型烟草的烟碱含量差异明显, PMT、QPT、CYP82E5V2、A622 4个烟碱合成代谢途径的关键基因的相对表达量受烟草种质类型、生育期、表达部位的影响不同, 烟碱含量变化与CYP82E5V2、PMT、A622的相对表达量呈显著正相关关系, 与QPT基因的相对表达量相关性不显著。本文烟草生长发育过程中的烟碱含量变化规律, 可为生产上降低烟碱含量措施的选择时期起到指导作用; 而烟碱含量与相关基因表达的相关性结果, 可为从分子水平阐述烟碱合成代谢机理和生产上利用生物技术降低或提高烟草的烟碱含量提供一定的理论参考。
(责任编辑 侯春晓)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|