{kind=link}
{kind=link}
番茄果实中主要风味物质研究进展
[王同林1  , 叶红霞
, 叶红霞2 , 郑积荣1, * , 李明3 ]
, 叶红霞, 李明|
|


作者简介:王同林(1992—),男,山东德州人,硕士,主要从事番茄育种工作。E-mail:757517470@qq.com
风味是番茄一个重要的品质指标,近年来受到人们的广泛关注。番茄的风味包括甜味、酸味、香味等,与果实中的可溶性糖含量、有机酸含量、挥发性芳香物质种类及含量有关。本文从影响番茄风味的主要组成成分,以及这些成分对番茄风味的贡献入手,对番茄果实主要风味贡献物质及其生物合成途径和其中涉及的主要酶展开讨论,对这些物质形成的遗传机理方面的国内外相关研究进行综合论述,就番茄果实风味研究存在的问题进行了分析,并对番茄品质研究前景进行展望,以期为今后番茄风味品质研究和育种工作提供参考。
Flavor is an important quality index of tomato, which has been widely concerned in recent years. The flavor of tomato includes sweetness, acidity and fragrance, which are related to the content of soluble sugar, organic acid and volatile aromatic compounds. In this paper, starting from the important components that affect the flavor of tomato and their contribution to the flavor of tomato, the main flavor contributing substances and their biosynthesis pathway of tomato fruit and the main enzymes involved were discussed, and the international research on the genetic mechanism of these substances formation were comprehensively discussed. The existing problems in the study of tomato fruit flavor were analyzed and the prospect of tomato quality research was proposed in order to provide reference for tomato quality research and breeding in the future.
番茄(Solanum lycopersicum)起源于南美洲, 16世纪传入欧洲, 17世纪传入中国。番茄是喜温性蔬菜, 几乎适应所有气候, 在世界各地广泛栽培。全球番茄种植面积约480万hm2(FAO, 2016), 中国是世界上最大的番茄生产国家, 番茄种植面积约为110万hm2, 产量在5 000万t以上, 基本可以自给自足, 还有部分用于出口(中国农业部, 2016)。
番茄的果实品质由外观品质、风味品质、营养品质和加工贮藏品质构成[1]。其中, 风味品质包括甜味、酸味、香味等, 与果实中的可溶性糖、有机酸和挥发性芳香化合物相关。风味丧失会严重影响果实的感官品质, 减少人们对果实的喜好程度, 进而影响其经济价值。因此, 如何在不影响或较小影响果实商品品质的前提下改善果实的风味, 成为番茄栽培和品质育种上需要重点解决的课题之一。
通过栽培管理技术(水分、施肥元素、温度、光照、土壤盐分及外源激素处理), 以及改变贮藏条件与手段来改善果实风味是当下热点研究内容。近年来, 国内外学者在番茄品质性状的遗传规律与数量性状基因座(quantitative trait locus, QTL)定位方面做了大量研究, 从数量遗传学方面揭示了番茄风味的遗传特征。科学家们还通过基因组测序、转录组与代谢组关联分析, 深层次阐明了番茄果实相关发育的进化历程、番茄育种过程的基因驯化及代谢物演变过程, 明确了番茄风味物质基础及遗传基础[2, 3, 4, 5], 这些工作的完成也为今后番茄风味改良育种奠定了基础。本文从番茄果实风味物质组成种类、生物合成代谢途径及其遗传机理等方面进行阐述, 提出了目前番茄品质研究中存在的问题并对未来发展前景进行了展望。
番茄果实中的可溶性糖类型因番茄不同种属而异, 普通番茄(Solanum lycopersicoides)和醋栗番茄(S. pimpinellifolium)中主要为葡萄糖和果糖; 秘鲁番茄(S. peruvianum)、克梅留斯基番茄(S. chmielewskii)和多毛番茄(S. habrochaites)中主要为蔗糖[6, 7]。除此之外, 番茄中还有微量的龙胆二糖、密二糖、甘露糖、尼日二糖等。不同类型的糖对番茄甜味的贡献也不同, 果糖甜度高, 葡萄糖甜度低。果糖、蔗糖和葡萄糖的甜度比为220: 145: 100[8]。
番茄果实内的可溶性糖主要由光合产物同化运输而来, Powell等[9]的研究表明, 番茄果实20%的光合产物来自果实自身的光合作用, 80%来自叶片等源器官。蔗糖是番茄光合作用的主要同化产物, 也是番茄中光合产物转运的主要形式。在番茄果实的发育期, 光合产物以蔗糖形式, 通过共质体和质外体途径经由韧皮部从叶片中直接转运到果柄甚至果实中, 进而在果实中各种酶的作用下进行代谢(图1)[10]。
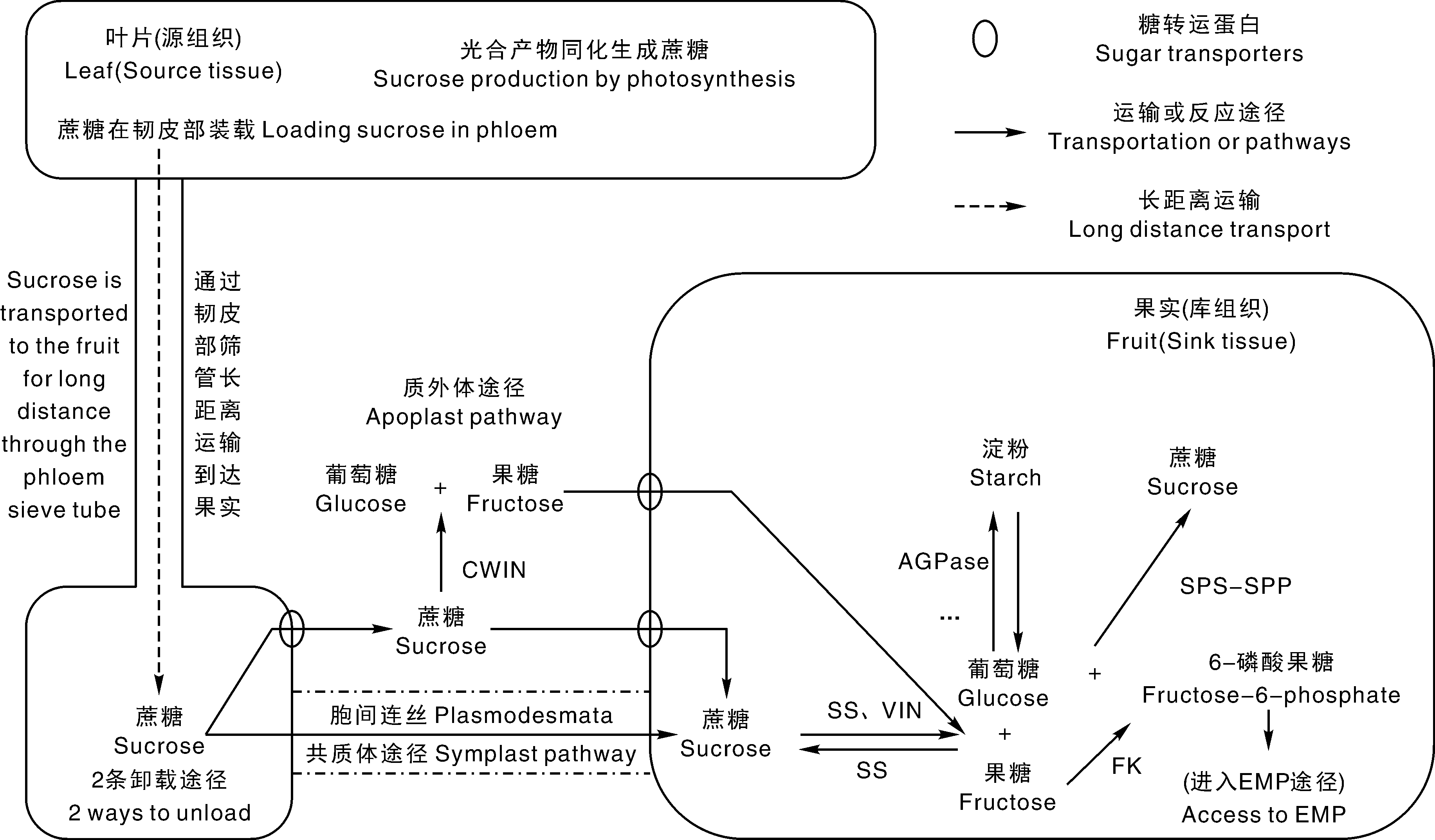 | 图1 番茄中蔗糖转运途径及可溶性糖的代谢途径 CWIN, 细胞壁转化酶; SS, 蔗糖合成酶; VIN, 液泡蔗糖转化酶; AGPase, 腺苷二磷酸葡萄糖焦磷酸化酶; FK, 果糖激酶; SPS, 蔗糖磷酸合成酶; SPP, 磷酸蔗糖酶。Fig.1 Sucrose transport pathway and soluble sugar metabolism pathway in tomato CWIN, Cell wall invertase; SS, Sucrose synthetase; VIN, Vacuolar invertase; AGPase, ADP-Glc pyrophosphorylase; FK, Fructokinase; SPS, Sucrose phosphate synthase; SPP, Sucrose phosphatase. |
GLK1和GLK2是2个功能相似的MYB类转录因子, 分别调控叶片和果实中叶绿体发育与叶绿素含量, 对番茄的光合作用有重要的调节作用。GLK2是U基因表达产物, 能够提高调控光合作用的基因转录丰度, 并增加未成熟果实中的淀粉含量, 进而使成熟果实中葡萄糖和果糖的含量提高40%左右; 另一方面, 在其调控作用下, 靠近番茄果柄的果实部分呈深绿色(绿果肩), 转色较慢, 影响了番茄的成熟及其外观品质[9], 因此, 当育种者对有利于番茄均匀成熟的无绿果肩性状进行选育时, 会导致番茄果实内的光合产物流入量降低。
蔗糖在叶片等源器官韧皮部进行装载, 经筛管长距离运输到果实后, 向果实库细胞进行卸载。蔗糖卸载存在2条途径:共质体途径和质外体途径。在果实发育前期, 果实内部的筛管、韧皮部与周围库细胞之间存在大量的胞间连丝, 源器官主要通过共质体途径向番茄果实运输蔗糖, 并且此时进入的蔗糖多代谢合成为淀粉贮藏在果实中, 但到果实发育后期, 由于果糖和葡萄糖的积累, 渗透势提高, 为了保持浓度梯度, 防止蔗糖回流到筛管, 这些胞间连丝开始退化, 蔗糖卸载途径逐步变为以质外体途径为主(图1)[11]。
番茄中的可溶性糖在质外体进入库细胞是通过糖转运蛋白实现的, 目前大部分的糖转运蛋白都是消耗能量来实现糖的转运, 因此, 会使果实发育受到一定影响。这可能是风味好的番茄品种果型较小、产量较低的原因之一。SWEETs蛋白转运糖不需要消耗能量, 为糖转运机制提供了新思路, 但其对糖的低亲和性, 使其具有局限性, 在糖含量顺浓度梯度条件下具有高效率, 在逆浓度梯度下的转运效果不佳[12]。
谷婧玥[13]利用高糖和低糖的2个番茄高代自交系构建6世代, 对分离世代果实中的可溶性糖含量进行测定分析发现, 番茄中的可溶性糖含量性状属于数量遗传中加性— 显性遗传。任婧[14]的研究表明, 番茄中的可溶性糖含量更符合一对加性— 显性主基因+加性— 显性— 上位性多基因遗传模型(MX1-AD-ADI), 对可溶性糖进行进一步遗传分析表明, 番茄果实可溶性糖含量的遗传效应以多基因效应为主, 环境对其影响相对较小。Tieman等[4]的报道指出果实质量和可溶性糖含量呈强负相关。
番茄果实中与可溶性糖代谢相关的酶包括蔗糖合成酶、蔗糖磷酸合成酶、磷酸蔗糖酶、蔗糖转化酶、腺苷二磷酸葡萄糖焦磷酸化酶、果糖激酶等。
1.4.1 蔗糖合成酶
蔗糖合成酶(sucrose synthetase, SS, EC 2.4.1.13)催化的是可逆反应, 既可以催化葡萄糖和果糖结合形成蔗糖(pH 8.0~9.5), 也可以催化蔗糖水解成果糖和葡萄糖(pH 5.5~6.0), 催化作用偏向水解方向。SS由TOMSSF家族基因表达合成, 在果实发育前期, SS活性与果实库强正相关, 控制着蔗糖向果实的转运量[11]。
1.4.2 蔗糖磷酸合成酶
蔗糖磷酸合成酶(sucrose phosphate synthase, SPS, EC 2.4.1.14)催化葡萄糖和果糖-6-磷酸结合形成磷酸蔗糖, 为可逆反应, 而磷酸蔗糖酶(sucrose phosphatase, SPP, EC 3.1.3.24)催化磷酸蔗糖水解成蔗糖, 使SPS催化的反应基本不可逆, 所以SPS-SPP系统是蔗糖合成的主要途径。SPS最适pH为7.0, 在光合器官和非光合器官都存在, SPS活性的提高, 可以增加叶片中蔗糖的合成量及果实中蔗糖的积累量[11]。
1.4.3 蔗糖转化酶
蔗糖转化酶(invertase, INV, EC 3.2.1.26)能够不可逆地催化蔗糖的水解反应, 生成葡萄糖和果糖, 是番茄中蔗糖水解主要的途径; 根据最适pH值, INV分为酸性蔗糖转化酶(acid invertase, AI)和中性蔗糖转化酶(neutral invertase, NI), AI的最适pH为3~5, 其活性占到总转化酶的95%, 是番茄果实中主要的蔗糖转化酶。而根据亚细胞定位, 可以将INV分为液泡蔗糖转化酶(vacuolar invertase, VIN)和细胞壁蔗糖转化酶(cell wall invertase, CWIN)。VIN存在于液泡内, 可以调节液泡中的蔗糖代谢; CWIN位于细胞间隙并结合在细胞壁上, 主要负责水解从韧皮部向番茄果实质外体卸载的蔗糖。Chetelat等[15]利用果实中积累蔗糖的克梅留斯基番茄与果实中积累葡萄糖和果糖的普通番茄杂交, 发现F1代果实均不积累蔗糖, 由此确定蔗糖的积累由隐性基因控制, 并在第3条染色体上定位到了sucr(sucrose accumulator)这一位点, 而sucr正是编码VIN基因(SUCR)的等位基因。Chetelat等[16]利用分子标记, 将该位点导入普通番茄中, 发现可以显著提高成熟果实中的蔗糖水平, 而且可溶性固形物含量也有所增加。也有学者将反义酸性转化酶基因SUCR导入普通番茄中, 得到的植株果实蔗糖浓度有明显的提高, 但总的可溶性糖含量并没有显著变化[17], 表明该基因仅对可溶性糖组分产生影响, 并不能提高果实内的可溶性糖含量; 另外, Levin等[18]在第4染色体上定位到一对调控果实内葡萄糖和果糖比例等位基因(FgrH/Fgr), 并表明FgrH对Fgr成不完全显性, FgrH的表达会使果糖占比增高, 但对总糖含量无明显影响, 该位点可能和蔗糖转化酶存在一定关系。Fridman等[19, 20]在与绿果潘那利番茄组配的渐渗系群体定位到一个可以提高番茄果实可溶性固形物含量的QTL位点Brix9-2-5(Lin5), 其编码CWIN, 并表明该位点可以提高番茄果实库强, 进而显著提高果实葡萄糖和果糖含量。Tieman等[4]通过代谢组与全基因组关联分析技术(metabolome genome-wide-association study, mGWAS)和代谢组与数量性状基因座定位技术(metabolome quantitative trait locus, mQTL)定位到9个与糖类物质相关的基因位点, 明确了位于9号和11号染色体上的2个调控番茄中的可溶性糖含量主效基因位点Lin5和SSC.11.1。
1.4.4 腺苷二磷酸葡萄糖焦磷酸化酶
腺苷二磷酸葡萄糖焦磷酸化酶(ADP-Glc pyrophosphorylase, AGPase, EC 2.7.7.21)催化番茄中淀粉合成的第一步, 是淀粉合成限速酶。Petreikov等[21]在多毛番茄中发现一个编码AGPase大亚基的基因(AGPL1), 研究发现, 该基因是通过提高未成熟果实中瞬时淀粉(过渡型淀粉)含量, 进而增加番茄果实中的可溶性糖含量。天然合成的AGPase是由2个大亚基和2个小亚基构成的异四聚体。早期学者认为, 小亚基具有催化活性, 大亚基只有调节作用, 后来大亚基被证实也具有一定的催化活性; 每个亚基由不同的基因控制合成, 这些基因在番茄植物体中表达的时空分布, 体现出其对库源器官中可溶性糖的调控作用[21]。
1.4.5 果糖激酶
果糖激酶(fructokinase, FK, EC 2.7.1.4)是果糖代谢的关键酶, 不可逆催化果糖磷酸化形成6-磷酸果糖, 磷酸化的果糖可以提高韧皮部和库细胞的蔗糖梯度, 帮助建立组织的库强[22]。6-磷酸果糖可以进一步磷酸化形成2, 6-磷酸果糖参与糖酵解途径, 也可以经SPS-SPP催化形成蔗糖; 果糖激酶还可以作为植物的己糖感受器和信号分子。因此, 果糖激酶对果实生长发育与可溶性糖含量有重要的调节作用。
番茄中的酸味由有机酸含量决定, 有机酸通常是指分子结构中含有羧基(-COOH)的酸性有机化合物。番茄果实中有机酸主要为柠檬酸和苹果酸, 且以柠檬酸为主, 另外番茄果实中还含有微量的草酸、莽草酸、反丁烯二酸、琥珀酸、酒石酸、抗坏血酸等。
在加工番茄中, 酸度越高越有利于增加番茄加工制品的保存时间[23]。在口感上, 有机酸能增强水果消费过程中对糖的感知, 其中柠檬酸比苹果酸更酸, 且对番茄风味的改良效果更好[4]。Da Cruz Centeno等[24]通过转反义延胡索酸酶基因和苹果酸脱氢酶基因来改变番茄果实中苹果酸浓度, 进而测定果实中淀粉和糖的含量, 得出番茄中的苹果酸浓度与过渡型淀粉含量(主要由AGPase调控)和可溶性固形物含量呈强负相关, 并表明番茄果实中的苹果酸含量虽不能影响果实的成熟度, 但与果实采后软化过程密切相关。
有机酸是渗透势形成的主要物质, 对发育期果实的膨胀有重要作用[25]。在果实发育前期, 果实中的有机酸总含量随时间推移不断增加, 而在果实成熟的后期伴随糖含量的迅速提高逐步减少, 而在最后的成熟阶段, 有机酸含量又再次增加[26]。成熟番茄的裂果现象也跟发育后期果实内的有机酸含量过高有一定关系[25]。
番茄果实中有机酸来源主要依赖于线粒体中的三羧酸循环(TCA)。TCA是三大营养素(糖、脂肪酸、氨基酸)的最终氧化途径, 为机体供能的同时也是3个物质的代谢联系枢纽。苹果酸脱氢产生的草酰乙酸与乙酰CoA在柠檬酸合酶的催化作用下生成柠檬酸, 柠檬酸经过多种酶逐步催化, 再次生成苹果酸形成循环(图2)。TCA中的多种有机酸处于动态平衡中, 途径中的多种中间产物还会参与光合作用、呼吸作用与多种芳香性挥发物质的代谢等被消耗[27], 此时磷酸烯醇式丙酮酸及丙酮酸在羧化酶的催化作用下生成草酰乙酸, 为TCA补充底物, 保持循环的顺利进行[28]。另外, 不同形态氮素的同化也会影响番茄果实中有机酸的代谢。根部吸收
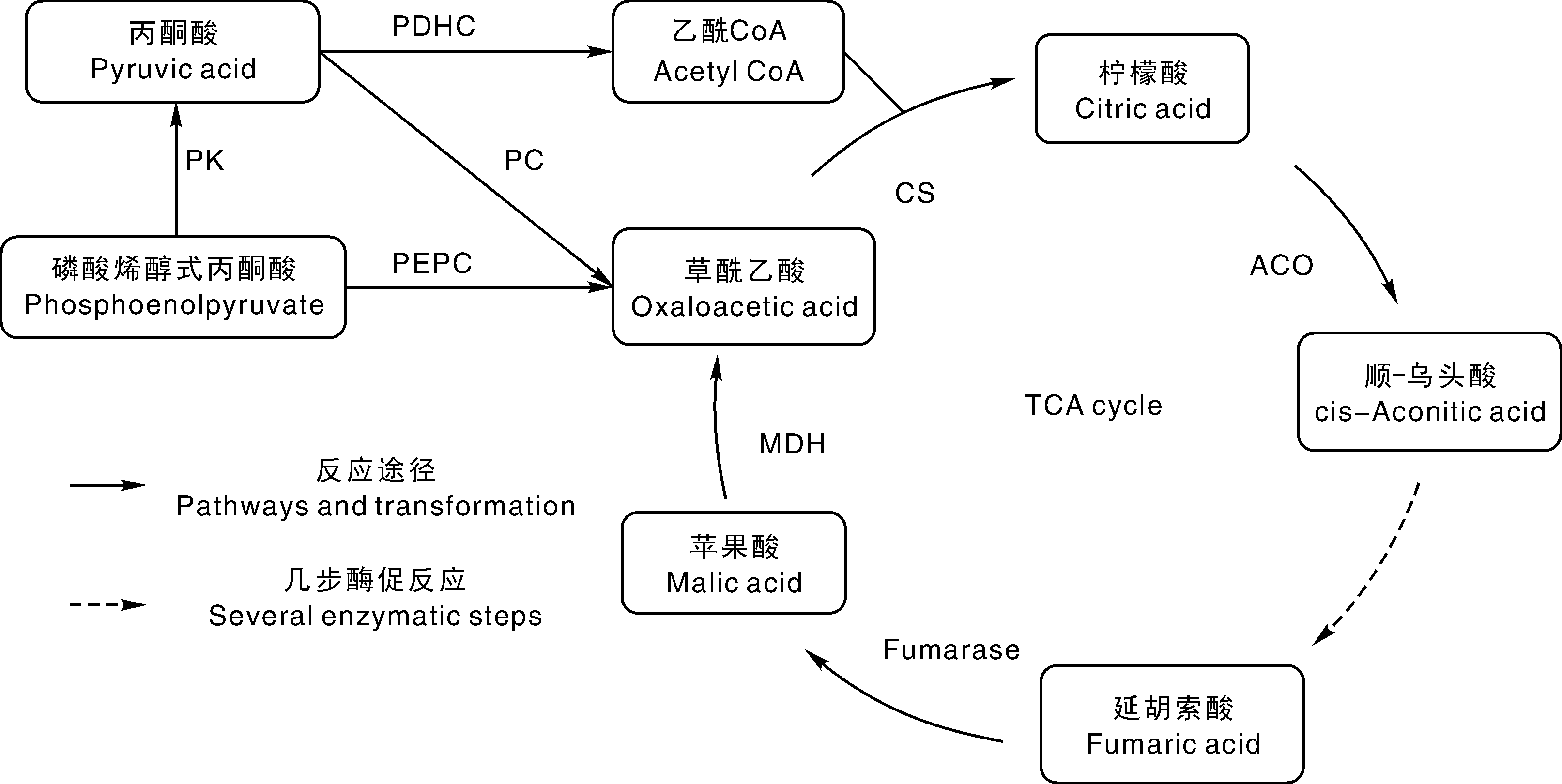 | 图2 三羧酸循环 PDHC, 丙酮酸脱氢酶; PK, 丙酮酸激酶; PC, 丙酮酸羧化酶; PEPC, 磷酸烯醇式丙酮酸羧化酶; CS, 柠檬酸合酶; ACO, 乌头酸水合酶; MDH, 苹果酸脱氢酶。Fig.2 Tricarboxylic acid cycle PDHC, Pyruvate dehydrogenase; PK, Pyruvate kinase; PC, Pyruvate carboxylase; PEPC, Phosphoenolpyruvate carboxylase; CS, Citrate synthase; ACO, Aconitase; MDH, Malate dehydrogenase. |
果实中多个有机酸代谢酶与有机酸水平相关(图2), Guillet等[29]对2个番茄中分别位于12号和7号染色体上调控PEPC的位点LYCesPpc1和LYCesPpc2进行研究发现, 在果实生长初期, LYCesPpc2的表达与果实内的柠檬酸和苹果酸一致增加, 而且表明PEPC不受胞质pH值降低及苹果酸浓度升高抑制, 推测PEPC可能在果实发育前期的有机酸合成和果实膨大起着重要作用。CS调控柠檬酸合成, ACO调控柠檬酸的降解, 一般认为CS和ACO催化反应的平衡调节柠檬酸含量, 但实际其机理非常复杂, 因为柠檬酸在线粒体中合成, 而在液泡中贮藏或积累[30]。Morgan等[31]通过对番茄中编码ACO基因(SlAco3b)的转反义基因株系进行分析发现, ACO活性降低, 而果实成熟时柠檬酸及苹果酸含量显著提高, 有机酸总含量提高了近50%, 证明ACO活性的降低有利于番茄果实内有机酸的积累。番茄中苹果酸含量由苹果酸的合成(fumarase)、降解(MDH)和细胞内转运(苹果酸转运蛋白)3个过程决定[32]。Ye等[33]利用mGWS等技术鉴定了位于6号染色体上的苹果酸积累的数量遗传位点TFM6, 其与Al-ACTIVATED MALATE TRANSPORTER9(Sl-ALMT9)相对应。Sl-ALMT9编码一种铝激活的苹果酸转运蛋白, 其表达有利于增加果实和根部的苹果酸含量, 根部的苹果酸阴离子可以螯合Al3+, 从而减轻铝胁迫, 进而提高了番茄风味[33]。
番茄的香味主要来自果实中的挥发性芳香化合物。在番茄中挥发性物质多种多样, 目前已经有400多种挥发性芳香化合物被鉴定出来, 其中只有几十种物质对番茄芳香存在贡献, 主要包括醇类、醛类、酮类、萜类和酯类物质以及含硫化合物等[4]。Buttery等[37]在研究中指出, 将顺-3-己烯醛、顺-3-己烯醇、己醛、1-戊烯-3-酮、3-甲基丁醇、反-2-己烯醛、6-甲基-5-庚烯-2-酮、2-异丁基硫咪唑、甲基水杨酸、β -紫罗兰酮10种物质在适当的浓度下混合, 就可以获得新鲜成熟番茄的芳香味道。
番茄的香味感知方式有2种途径, 一种是果实成熟时发出的香气直接进入鼻腔被嗅觉感知, 另一种是咀嚼果肉时发散出来的香气通过口腔进入鼻腔被感知, 2种挥发物传递方式在大脑中的感知方式不同[38]。食品风味化学定义每一种芳香物质都有固定阈值浓度, 只有超过该阈值, 才能被人们嗅觉所感知, 而用含量与该阈值浓度比值的对数来衡量某种物质对香气的作用效果, 对数值大于0, 则表明对风味的贡献较大。而Baldwin等[39]认为, 一些挥发性物质阈值单位对数即使为负数, 仍会对番茄的整体风味有所贡献。
番茄中芳香物质合成途径按前体物质大致可分为4类:以脂肪酸为前体的合成途径、以类胡萝卜素为前体的合成途径、以苯丙氨酸为前体的合成途径和以支链氨基酸为前体的合成途径[40]。目前已经有很多控制果实挥发性芳香化合物合成的相关酶基因被克隆出来。
3.2.1 以脂肪酸为前体的合成途径
脂肪酸前体物质主要是指亚油酸和亚麻酸, 主要有三大主要酶系统参与脂肪酸向挥发性芳香物质的代谢:脂氧合酶系统(LOX)、脂氢过氧化物裂解酶系统(HPL)和氧化还原酶系统(ADH)[41]。酯氧合酶C(lipoxygenase C, LOXC, EC 1.13.11.12)催化亚油酸和亚麻酸形成脂氢过氧化物, 而在13-脂氢过氧化物裂解酶(hydroperoxide lyase, 13-HPL, EC 4.1.2.-)进一步催化作用下形成短链挥发性C6醛类物质(己醛、己烯醛等), 乙醇脱氢酶(alcohol dehydrogenase, ADH, EC1.1.1.1)催化C6醛类物质与C6醇类物质(己醇、己烯醇等)相互转化。樱桃番茄中脂肪酸含量是普通番茄中的2倍以上, 这可能是樱桃番茄比普通番茄风味更好的原因之一[42]。该合成途径中主要酶基因Tomloxc[43]、13-HPL和ADH2已经被相继克隆出来[44]。
3.2.2 以苯丙氨酸为前体的合成途径
苯丙氨酸是4种重要挥发物质(苯乙醛、2-苯乙醇、1-硝基-2-苯乙烷、2-苯乙腈)的前体。LeAADC编码氨基酸脱羧酶(amino acid decarboxylases, AADCs, EC4.1.1.-), 催化苯丙氨酸脱羧生成苯乙胺, 是苯丙氨酸向4种挥发性物质代谢的第一步, AADCs也是该代谢途径中的限速酶; 苯乙胺进一步被催化合成2-苯乙醛、2-苯乙腈、1-硝基-2-苯乙烷。LePARs编码苯乙醛还原酶(phenylacetaldehyde reductases, PARs, EC1.1.1.2), 催化2-苯乙醛还原为2-苯乙醇。此外, 苯丙氨酸也可先经苯丙氨酸解氨酶(phenylalanine ammonia lyase, PAL, EC 4.3.1.5)催化形成反式肉桂酸(trans-cinnamic acid), 进一步催化形成水杨酸、儿茶酚等。水杨酸羧基甲基转移酶(salicylic acid carboxylmethyltransferase, SAMT1, EC2.1.1.-)、儿茶酚氧位甲基转移酶(catechol O-methyl transferase, CTOMT1, EC2.1.1.6)分别催化水杨酸和儿茶酚甲基化生成水杨酸甲酯及愈创木酚[45]。目前该合成途径中的LeAADC[46]、LePAR1和LePAR2[47], SlSAMT1[48]、CTOMT1[49]等基因已被克隆出来。
3.2.3 以支链氨基酸为前体的合成途径
支链氨基酸前体物质主要是指亮氨酸、异亮氨酸与缬氨酸等。支链氨基酸氨基转移酶(branched-chain amino acid aminotransferases, BCATs, EC 2.6.2.42)催化支链氨基酸脱氨形成酮酸, 进一步脱氢生成醇类、酮类、酯类物质, 醇类经氧化生成醛类, 如2-甲基丁醇、2-甲基丁醛、3-甲基丁醇、3-甲基丁醛、乙酸异丁酯、异戊腈和2-异丁基噻唑。其中2-异丁基噻唑为番茄所特有, 而3-甲基丁醇能够增加人们对甜度的感觉[4]。目前在番茄中已有SlBCAT1-6共6个支链氨基酸转氨酶基因被克隆出来[44]。
3.2.4 以类胡萝卜素为前体的合成途径
番茄果实中的类胡萝卜素主要包括:番茄红素和β -胡萝卜素等, LeCCD1编码类胡萝卜素裂解双加氧酶(carotenoid cleavage dioxygenases, CCDs, EC1.13.11), 对类胡萝卜素进行裂解产生一些挥发性萜类化合物, 例如香叶醛、橙花醛、6-甲基-5-庚烯-2-酮、香叶基丙酮、假紫罗酮、β -紫罗兰酮和β -大马酮, 它们均具有花香果香, 对番茄香气有重要贡献[44]。各类挥发性芳香化合物由不同色素特异合成, 6-甲基-5庚烯-2-酮、香叶醛、橙花醛、假紫罗酮等由番茄红素裂解合成; 香叶基丙酮、β -紫罗兰酮、β -环柠檬醛等由β -胡萝卜素裂解合成; α -紫罗兰酮由δ -胡萝卜素裂解合成[44, 49]。目前, 该途径已有2个类胡萝卜素裂解双加氧酶基因被克隆:LeCCD1A和LeCCD1B[50]。
番茄果实的可溶性糖、有机酸、挥发性芳香化合物等风味物质都是由进入果实的蔗糖代谢而来。蔗糖进入果实后在蔗糖转化酶等作用下分解成葡萄糖和果糖, 可以进一步磷酸化进入糖酵解途径, 也可在酶的催化作用下合成淀粉等以保持库源蔗糖浓度梯度, 糖酵解产物丙酮酸又可以进入三羧酸循环, 参与柠檬酸和苹果酸的代谢, 在此循环中丙酮酸被进一步氧化分解成CO2和H2O, 产生大量能量参与果实内的各种生命活动。而糖酵解途径与莽草酸途径、甲羟戊酸途径等相交叉促进了类胡萝卜素、苯丙氨酸、亚油酸、亚麻酸及支链氨基酸等的合成[44]。其中有机酸在番茄果实发育过程中的积累、代谢与转运, 引起质体内pH值变化, 进而抑制或激活风味物质代谢过程中相关酶的活性, 进一步影响风味物质的代谢。因此, 光合同化物的流入是番茄果实中风味物质形成的基础, 而番茄风味物质种类与含量的调控则依赖于果实中各类酶系统之间的相互作用。
近十多年, 番茄新品种选育一直以硬度、产量、抗病性、果实大小和外观等性状为番茄的育种主要评价指标, 导致番茄的风味逐步下降:大果型番茄的选育使一些低糖位点以搭便车效应进入果实[4]; 均匀成熟(无绿果肩)的种质筛选, 使得一种转录因子失活, 导致番茄果实中可溶性固形物和类胡萝卜素减少[9]; 为引入野生番茄的抗病基因, 导致一些不良性状片段渗入, 降低了果实中风味相关的代谢产物[5]; 栽培番茄的提早采摘及贮运过程中的冷藏对番茄的风味也有一定影响[34]。
糖酸比指果实中可溶性总糖与可滴定总酸的比值, 是衡量口感的参考指标之一。谭其猛[6]认为, 番茄合适的糖酸比为6.9~10.8。也有研究指出, 番茄果实合适的糖酸比为7: 10[52]。仅可溶性糖含量、有机酸含量、糖酸比与挥发性芳香化合物等生理指标并不能确切地反映果实的风味, 口感是番茄风味品质的直观表现, 而番茄的口感跟果肉质地(硬度和肉质)也有一定的相关性, 硬度与口感品质一般呈负相关, 即硬度越高口感越差, 在硬度方面已有不少基因位点被报道, 但肉质评价如多汁性、颗粒度等方面的研究报道相对较少, 其中的遗传机理尚不明晰。这与风味评价系统尚不完善有一定关系。随着人们对番茄风味的要求不断提高, 番茄食用方式也回归多元化, 无论是菜用、水果鲜食, 番茄的风味需求越来越高。笔者认为, 番茄风味评价系统的研究和完善, 有利于番茄品质育种的多元化发展; 育种者应该积极借鉴水果的风味评价标准, 完善番茄的风味评价, 结合分子标记技术, 加快优良种质的筛选和创新; 结合本地区气候、土壤条件, 综合种植者和消费者对品种、品质的需求, 有目的地研发或改良以开发出符合市场需求的番茄新品种。
(责任编辑 张韵)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|