Human societal development: is it an evolutionary transition in individuality?
1
2023
... 近年来,随着生活水平的提高,消费者在选购瓜果蔬菜产品过程中越来越注重产品的外观品质.果蔬产品的色泽不仅是反映其成熟度的关键因素,同时也是影响其外观品质的核心因素[1 ⇓ -3 ] .果蔬菜品的商业价值存在较强的地域性,不同的地区消费者的喜好不同,因此,根据不同的市场需求来改良果蔬的外观性质,尤其是果皮颜色,对提高果蔬的商业价值具有重要意义. ...
Reduced nitrogen fertilization from pre-flowering to pre-veraison alters phenolic profiles of Vitis vinifera L. Cv. Cabernet Gernischt wine of Yantai, China
1
2023
... 近年来,随着生活水平的提高,消费者在选购瓜果蔬菜产品过程中越来越注重产品的外观品质.果蔬产品的色泽不仅是反映其成熟度的关键因素,同时也是影响其外观品质的核心因素[1 ⇓ -3 ] .果蔬菜品的商业价值存在较强的地域性,不同的地区消费者的喜好不同,因此,根据不同的市场需求来改良果蔬的外观性质,尤其是果皮颜色,对提高果蔬的商业价值具有重要意义. ...
Edible gels with cranberry extract: evaluation of anthocyanin release kinetics
1
2023
... 近年来,随着生活水平的提高,消费者在选购瓜果蔬菜产品过程中越来越注重产品的外观品质.果蔬产品的色泽不仅是反映其成熟度的关键因素,同时也是影响其外观品质的核心因素[1 ⇓ -3 ] .果蔬菜品的商业价值存在较强的地域性,不同的地区消费者的喜好不同,因此,根据不同的市场需求来改良果蔬的外观性质,尤其是果皮颜色,对提高果蔬的商业价值具有重要意义. ...
Recent advances on the development and regulation of flower color in ornamental plants
2
2015
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
... [4 ,7 ].例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Carotenoid metabolism and regulation in horticultural crops
1
2015
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Characterization and functional identification of a novel plant 4, 5-extradiol dioxygenase involved in betalain pigment biosynthesis in Portulaca grandiflora
1
2004
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Expression pattern analysis of three R2R3-MYB transcription factors for the production of anthocyanin in different vegetative stages of Arabidopsis leaves
1
2021
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Comparative analysis of the effects of internal factors on the floral color of four Chrysanthemum cultivars of different colors
1
2022
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Antioxidant activity of nasunin, an anthocyanin in eggplant peels
1
2000
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
How genes paint lily flowers: regulation of colouration and pigmentation patterning
1
2013
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Anthocyanin accumulation in poinsettia leaves and its functional role in photo-oxidative stress
1
2020
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Molecular biology of anthocyanin accumulation in grape berries
1
2009
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Differential expressions of anthocyanin synthesis genes underlie flower color divergence in a sympatric Rhododendron sanguineum complex
1
2021
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Functional R2R3-MYB transcription factor NsMYB1, regulating anthocyanin biosynthesis, was relative to the fruit color differentiation in Nitraria sibirica Pall
1
2022
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Upregulation of the MYB2 transcription factor is associated with increased accumulation of anthocyanin in the leaves of Dendrobium bigibbum
1
2020
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Comparison of flower color with anthocyanin composition patterns in evergreen Azalea
1
2009
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples
3
2006
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions
1
2012
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Accumulation of anthocyanins in detached leaves of Kalanchoë blossfeldiana : relevance to the effect of methyl jasmonate on this process
1
2022
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Evaluating potential of leaf reflectance spectra to monitor plant genetic variation
1
2023
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Antioxidant activities, phenolic and β-carotene contents of sweet potato genotypes with varying flesh colours
2
2007
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
Purple sweet potato color attenuates oxidative stress and inflammatory response induced by D
1
2009
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Antimutagenicity of deacylated anthocyanins in purple-fleshed sweetpotato
1
2001
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
In vivo antioxidant, hypoglycemic, and anti-tumor activities of anthocyanin extracts from purple sweet potato
2
2013
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
... 花青素和多酚类化合物对肿瘤的特殊抑制作用是近年来植物提取物治疗或辅助治疗癌症的研究热点.Zhao等[24 ] 的研究表明,小鼠体内植入S180肿瘤细胞后体重降低,然而紫甘薯花青素能够通过提高谷胱甘肽过氧化物酶和超氧化物歧化酶水平,增强抗氧化能力,丙二醛水平降低,从而抑制小鼠体内肿瘤生长;另一项关于紫甘薯的研究表明,花青素通过诱导细胞凋亡抑制膀胱癌BIU87细胞的增殖,且具有剂量依赖性[150 ] .此外,花青素对小鼠肿瘤生长的抑制率与花青素浓度成正比.槲皮素可抑制子宫肿瘤细胞的增殖,且其治疗效果优于其他活性物质.从葡萄中提取的花青素对乳腺癌MCF-7细胞的抗肿瘤作用优于纯化的花青素[151 ] ,这与Yang等[152 ] 的研究结果一致.另外,在一定程度上,活性物质对体外肿瘤增殖的抑制能力与抗氧化活性并不呈正相关[152 ] . ...
Anti-aging effects and mechanisms of anthocyanins and their intestinal microflora metabolites
1
2022
... 植物器官的各种颜色通常是由特定的类黄酮、类胡萝卜素和生物碱的种类和积累量的差异引起的[4 ⇓ -6 ] .花青素是最重要的类黄酮化合物之一,普遍存在于许多植物和水果中,对植物器官的色素沉着起着至关重要的作用[4 ,7 ] .例如,花青素显著影响紫红色菊花花瓣、深紫色茄子果实和粉红色百合花朵[8 ⇓ -10 ] .一品红红叶和葡萄红黑莓的颜色均与花青素的积累有显著的相关性[11 -12 ] .先前的研究表明,类黄酮生物合成途径中基因表达水平的变化导致植物器官中花青素大量积累,从而导致颜色多态性[13 ] .由于菠萝成熟果实中AcHOX 21和AcMYB 12相对表达水平下调,花青素含量减少,以及内源茉莉酸、赤霉素酸和生长素水平的波动,导致菠萝果皮因花青素介导的变色而变色.NsMYB 1基因促进西伯利亚白刺黑色果实花青素的积累[14 ] .DbMYB 2的瞬时过表达显著促进烟叶花青素积累[15 ] .紫色杜鹃花花朵中可以检测到各种各样的花青素,而白色花瓣中则没有检测到花青素.此外,还发现花瓣颜色较深的原因是其胚和种皮中花青素大量积累[16 ] .这些研究结果一致表明,颜色的增强与花青素水平的提高密切相关.阳光可以促进植物花青素的吸收,尤其是在苹果和葡萄的果皮中,缺乏光线则会产生相反的效果[17 -18 ] .花青素相对含量的高低可以明显地反映植物的生长生理状态,还可以显著提高植物非生物胁迫抗性[19 -20 ] .此外,花青素作为一种食品添加剂,具有多种生物活性,包括抗氧化性[21 ] 、抗炎症性[22 ] 、抗突变性和抗肿瘤性[23 -24 ] ,并在全国得到了广泛的应用.医学家利用花青素的强抗氧化性治疗人体癌症,发现有显著的效果[25 ] . ...
Editorial: advances in anthocyanins: sources, preparation, analysis methods, bioavailability, physiochemical properties, and structural features
1
2023
... 目前,已在自然界鉴定出6 000多种花青素,其中92%由天竺葵色素(Pg)、矢车菊素(Cy)、锦葵色素(Mv)、飞燕草色素(DP)、芍药色素(Pn)、牵牛花色素(Pt)等色素及其衍生物组成(表1 ).除了一系列酶和转录因子调控花青素合成以外,外界因素也发挥重要作用.木糖(C5 H10 O5 )、阿拉伯糖(C5 H10 O5 )、葡萄糖(C6 H12 O6 )、鼠李糖(C6 H12 O5 )等与花青素结合,通过糖基化、酰基化、甲基化等过程进一步进行修饰,在植物液泡中以糖苷的形式存在[26 ] .不同修饰方式及修饰程度对糖苷的种类和稳定性的影响均不相同,对糖苷种类的区分主要在于糖苷C骨架结构中的羟基数目、甲基化与酰基化的位置和种类数目(图1 ). ...
Identification and characterisation of anthocyanins from wild mulberry (Morus nigra L.) growing in Brazil
1
2007
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Determination of flavonoids and carotenoids and their contributions to various colors of rose cultivars (Rosa spp.)
2
2019
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
... [
28 ]
牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ] 10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Amazonian Melastomataceae blueberries: determination of phenolic content, nutritional composition, and antioxidant and anti-glycation activities
1
2022
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Optimization of ultrasound-assisted extraction of phenolic compounds and anthocyanins from blueberry (Vaccinium ashei ) wine pomace
1
2016
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Anthocyanin from Lycium ruthenicum Murr. in the Qaidam Basin alleviates ultraviolet-induced apoptosis of human skin fibroblasts by regulating the death receptor pathway
1
2022
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Anthocyanin-rich black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) and red cabbage (Brassica oleracea ) extracts incorporated biosensor for colorimetric detection of Helicobacter pylori with color image processing
1
2023
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Nanoencapsulation of anthocyanins from red cabbage (Brassica oleracea L. var. Capitata f. rubra ) through coacervation of whey protein isolate and apple high methoxyl pectin
1
2023
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Anthocyanins identification and transcriptional regulation of anthocyanin biosynthesis in purple Brassica napus
1
2022
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Molecular characterization of Prunus lusitanica L. fruit extracts and their health-promoting potential in inflammation, diabetes, and neurodegenerative diseases
1
2023
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
Insights into the genes involved in ABA biosynthesis and perception during development and ripening of the Chilean strawberry fruit
1
2023
... Six kinds of anthocyanins and their substituted chemical group
Table 1 花青素种类 R1碳位官能团 R2碳位官能团 物种 参考文献 天竺葵色素 H H 桑葚Morus alba L.Rosa spp. [27 -28 ] 矢车菊色素Cyanidin (Cy) OH H 蓝莓Vaccinium spp. [29 ] 飞燕草色素 OH OH 黑枸杞Lycium ruthenicum Murr.Vaccinium spp. [30 -31 ] 芍药花色 OMe H 玫瑰Rosa rugosa Daucus carota L. [32 ]28 ] 牵牛花色素 OMe OH 马铃薯Solanum tuberosum Lycium ruthenicum Murr. [33 -34 ] 锦葵色素 OMe OMe 葡萄Vitis vinifera L.Vaccinium corymbosum [35 -36 ]
10.3969/j.issn.1004-1524.20230660.F0001 图1 花青素的结构 Structure of anthocyanins Fig.1 ![]()
在近10年的研究中,研究者对花青素的生物学功能进行了广泛的研究,对花青素的研究重点转向外源刺激信号对花青素生物合成的影响及其调控机制,如光照、温度、糖和外源调节物质(5-氨基乙酰丙酸ALA 、脱落酸ABA、乙烯Eth、褪黑素MT)等,但对花青素全面概述的报道较少.因此,本文就近年来对花青素的生物合成、外界影响因素、相关基因表达、转录因子调控机制及其生物活性和检测方法进行了系统的阐述,为今后花青素研究提供理论支持. ...
A wheat R2R3-MYB protein PURPLE PLANT1 (TaPL1) functions as a positive regulator of anthocyanin biosynthesis
2
2016
... 由苯丙氨酸到花色苷的转化过程即生成花青素的过程(图2 ).苯丙氨酸依次经苯丙氨酸裂解酶(PAL)、肉桂酸羟化酶(C4H)、香豆酰COA连接酶(4CL)、查尔酮合成酶(CHS)催化生成查尔酮(chalcone).查尔酮经查尔酮异构酶(CHI)催化生成柚皮素(naringenin),再经黄烷酮3-羟化酶(F3H)作用转化生成香橙素(dihydrokaempferol),又名二氢黄酮醇.香橙素之后分为两条支路.其一,依次经二氢黄酮醇-4-还酶(DFR)和花青素合成酶(ANS)催化生成天竺葵素(pelargonidin);其二,依次经过类黄酮3'-羟化酶(F3'H)、二氢黄酮醇-4-还酶和花青素合成酶催化生成矢车菊素(cyanidin).之后,经过糖基化、丙二酰化等过程后形成稳定的花青素[37 ⇓ -39 ] . ...
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
Research progress of proanthocyanidins and anthocyanidins
1
2023
... 由苯丙氨酸到花色苷的转化过程即生成花青素的过程(图2 ).苯丙氨酸依次经苯丙氨酸裂解酶(PAL)、肉桂酸羟化酶(C4H)、香豆酰COA连接酶(4CL)、查尔酮合成酶(CHS)催化生成查尔酮(chalcone).查尔酮经查尔酮异构酶(CHI)催化生成柚皮素(naringenin),再经黄烷酮3-羟化酶(F3H)作用转化生成香橙素(dihydrokaempferol),又名二氢黄酮醇.香橙素之后分为两条支路.其一,依次经二氢黄酮醇-4-还酶(DFR)和花青素合成酶(ANS)催化生成天竺葵素(pelargonidin);其二,依次经过类黄酮3'-羟化酶(F3'H)、二氢黄酮醇-4-还酶和花青素合成酶催化生成矢车菊素(cyanidin).之后,经过糖基化、丙二酰化等过程后形成稳定的花青素[37 ⇓ -39 ] . ...
ARF2 positively regulates flavonols and proanthocyanidins biosynthesis in Arabidopsis thaliana
1
2022
... 由苯丙氨酸到花色苷的转化过程即生成花青素的过程(图2 ).苯丙氨酸依次经苯丙氨酸裂解酶(PAL)、肉桂酸羟化酶(C4H)、香豆酰COA连接酶(4CL)、查尔酮合成酶(CHS)催化生成查尔酮(chalcone).查尔酮经查尔酮异构酶(CHI)催化生成柚皮素(naringenin),再经黄烷酮3-羟化酶(F3H)作用转化生成香橙素(dihydrokaempferol),又名二氢黄酮醇.香橙素之后分为两条支路.其一,依次经二氢黄酮醇-4-还酶(DFR)和花青素合成酶(ANS)催化生成天竺葵素(pelargonidin);其二,依次经过类黄酮3'-羟化酶(F3'H)、二氢黄酮醇-4-还酶和花青素合成酶催化生成矢车菊素(cyanidin).之后,经过糖基化、丙二酰化等过程后形成稳定的花青素[37 ⇓ -39 ] . ...
Research progress on anthocyanin-mediated regulation of ‘black’ phenotypes of plant organs
1
2023
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
SlMYB12 regulates flavonol synthesis in three different cherry tomato varieties
3
2018
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
... CHI 在花青素合成过程中也起着重要的催化作用,主要在查尔酮转化生成黄烷酮的过程中起催化作用.目前,已在番茄(Lycopersicon esculentum Mill.)[41 ] 、洋葱(Allium cepa )[56 ] 、大豆[Glycine max (Linn.) Merr.][57 ] 、菜豆(Phaseolus vulgaris L.)[58 ] 等植物中分离克隆出了CHI 基因,其表达对花青素以及类黄酮的积累有重要作用.将矮牵牛CHI 基因转入番茄中过度表达,转基因番茄果皮中类黄酮化合物含量增加78倍[41 ] .Morita等[59 ] 在日本牵牛花中鉴定出了一种花青素合成增强基因EFP ,发现敲除EFP 基因使牵牛花(Petunia hybrida )和斗牛花(Torenia hybrida )中花青素的含量显著降低,表现为花色褪去. ...
... [41 ].Morita等[59 ] 在日本牵牛花中鉴定出了一种花青素合成增强基因EFP ,发现敲除EFP 基因使牵牛花(Petunia hybrida )和斗牛花(Torenia hybrida )中花青素的含量显著降低,表现为花色褪去. ...
Anthocyanin biosynthesis and DNA methylation dynamics in sweet orange fruit[Citrus sinensis L. (osbeck)] under cold stress
1
2020
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
Gene transcript accumulation, tissue and subcellular localization of anthocyanidin synthase (ANS) in developing grape berries
1
2010
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
Identification and characterisation of F3GT1 and F3GGT1, two glycosyltransferases responsible for anthocyanin biosynthesis in red-fleshed kiwifruit (Actinidia chinensis )
1
2011
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
Introduction of apple ANR genes into tobacco inhibits expression of both CHI and DFR genes in flowers, leading to loss of anthocyanin
1
2012
... 多种因素共同调控着花青素的生物合成[40 ] .在结构基因中,合成通路上游PAL 、CHS 、CHI 、F 3H 等酶基因起到了重要调控作用,并且这些酶基因的表达参与果实发育进程[17 ,41 -42 ] .而在合成通路下游中发现DFR、ANS以及其他酰基化酶、糖基化酶等酶基因则主要调控果实果皮的颜色,且这些基因的高效表达及其酶活性与果皮颜色呈正相关关系[43 ⇓ -45 ] . ...
The phenylalanine ammonia lyase (PAL) gene family shows a gymnosperm-specific lineage
1
2012
... PAL是花青素合成初始反应的第一个关键限速酶,也是植物次生代谢的一个重要调控点,几乎存在于所有植物中[46 ] .对草莓喷施PAL抑制剂 L-α-氨基β苯基丙酸有效抑制了花青素的合成[47 ] .分别在未成熟和成熟苹果表皮上喷施乙烯,发现未成熟苹果中PAL活性与花青素积累水平增加,而成熟的苹果PAL活性与花青素积累水平无显著变化,说明乙烯通过激发PAL酶的活性,促进了苹果表皮花青素的积累[48 ] .对萝卜幼苗采用UV-B照射的处理,24 h后萝卜幼苗下胚轴花青素的积累水平迅速增加,发现萝卜幼苗花青素的积累与PAL活性的激活呈显著正相关[49 ] . ...
Cloning of FaPAL 6 gene from strawberry fruit and characterization of its expression and enzymatic activity in two cultivars with different anthocyanin accumulation
1
2011
... PAL是花青素合成初始反应的第一个关键限速酶,也是植物次生代谢的一个重要调控点,几乎存在于所有植物中[46 ] .对草莓喷施PAL抑制剂 L-α-氨基β苯基丙酸有效抑制了花青素的合成[47 ] .分别在未成熟和成熟苹果表皮上喷施乙烯,发现未成熟苹果中PAL活性与花青素积累水平增加,而成熟的苹果PAL活性与花青素积累水平无显著变化,说明乙烯通过激发PAL酶的活性,促进了苹果表皮花青素的积累[48 ] .对萝卜幼苗采用UV-B照射的处理,24 h后萝卜幼苗下胚轴花青素的积累水平迅速增加,发现萝卜幼苗花青素的积累与PAL活性的激活呈显著正相关[49 ] . ...
Anthocyanin accumulation in apple skin during ripening: regulation by ethylene and phenylalanine ammonia-lyase
1
1984
... PAL是花青素合成初始反应的第一个关键限速酶,也是植物次生代谢的一个重要调控点,几乎存在于所有植物中[46 ] .对草莓喷施PAL抑制剂 L-α-氨基β苯基丙酸有效抑制了花青素的合成[47 ] .分别在未成熟和成熟苹果表皮上喷施乙烯,发现未成熟苹果中PAL活性与花青素积累水平增加,而成熟的苹果PAL活性与花青素积累水平无显著变化,说明乙烯通过激发PAL酶的活性,促进了苹果表皮花青素的积累[48 ] .对萝卜幼苗采用UV-B照射的处理,24 h后萝卜幼苗下胚轴花青素的积累水平迅速增加,发现萝卜幼苗花青素的积累与PAL活性的激活呈显著正相关[49 ] . ...
Improving the anthocyanin accumulation of hypocotyls in radish sprouts by hemin-induced NO
1
2022
... PAL是花青素合成初始反应的第一个关键限速酶,也是植物次生代谢的一个重要调控点,几乎存在于所有植物中[46 ] .对草莓喷施PAL抑制剂 L-α-氨基β苯基丙酸有效抑制了花青素的合成[47 ] .分别在未成熟和成熟苹果表皮上喷施乙烯,发现未成熟苹果中PAL活性与花青素积累水平增加,而成熟的苹果PAL活性与花青素积累水平无显著变化,说明乙烯通过激发PAL酶的活性,促进了苹果表皮花青素的积累[48 ] .对萝卜幼苗采用UV-B照射的处理,24 h后萝卜幼苗下胚轴花青素的积累水平迅速增加,发现萝卜幼苗花青素的积累与PAL活性的激活呈显著正相关[49 ] . ...
The type Ⅲ polyketide synthase supergene family in plants: complex evolutionary history and functional divergence
1
2022
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
Chalcone isomerase cDNA cloning and mRNA induction by fungal elicitor, wounding and infection
1
1987
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
Structure and function of the chalcone synthase superfamily of plant type Ⅲ polyketide synthases
1
2010
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
甘薯查尔酮合成酶基因IbCHS1的克隆和表达分析
1
2018
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
甘薯查尔酮合成酶基因IbCHS1的克隆和表达分析
1
2018
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
Modification of flower color in Torenia (Torenia fournieri Lind.) by genetic transformation
1
2000
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
Flower color modulations of Torenia hybrida by downregulation of chalcone synthase genes with RNA interference
1
2004
... 4-香豆酸和丙二酰CoA经过CHS 催化生成查尔酮,为花青素后续合成和代谢构建基础碳架结构[50 ] .据报道,查尔酮合成酶是CHS 超基因家族的核心酶,自Mehdy等[51 ] 在1987年首次在豌豆中克隆并分离出CHS 基因后,目前,已从20多个植物和细菌中识别并克隆出了查尔酮超基因家族成员的基因序列[52 ] .在甘薯中,已克隆得到CHS编码基因IbCHS 1,经生物信息学分析发现IbCHS 1具有典型的查尔酮合成酶的结构特征,表达分析结果发现,其主要在甘薯果肉中表达,且表达量与甘薯中花青素含量呈显著正相关[53 ] .由于CHS 在花青素生物合成的上游代谢过程中发挥作用,研究发现,抑制查尔酮合成酶基因的表达时,培育出的花卉花色较淡.Aida等[54 ] 的研究发现,在夏堇中导入CHS 的反义基因后,其花色变浅;而Fukusaki等[55 ] 利用RNAi技术沉默了夏堇CHS 基因后,得到了花朵颜色为白色和灰白色的新品种. ...
Gold color in Onions (Allium cepa ): a natural mutation of the chalcone isomerase gene resulting in a premature stop codon
1
2004
... CHI 在花青素合成过程中也起着重要的催化作用,主要在查尔酮转化生成黄烷酮的过程中起催化作用.目前,已在番茄(Lycopersicon esculentum Mill.)[41 ] 、洋葱(Allium cepa )[56 ] 、大豆[Glycine max (Linn.) Merr.][57 ] 、菜豆(Phaseolus vulgaris L.)[58 ] 等植物中分离克隆出了CHI 基因,其表达对花青素以及类黄酮的积累有重要作用.将矮牵牛CHI 基因转入番茄中过度表达,转基因番茄果皮中类黄酮化合物含量增加78倍[41 ] .Morita等[59 ] 在日本牵牛花中鉴定出了一种花青素合成增强基因EFP ,发现敲除EFP 基因使牵牛花(Petunia hybrida )和斗牛花(Torenia hybrida )中花青素的含量显著降低,表现为花色褪去. ...
Genome-wide classification and evolutionary analysis reveal diverged patterns of Chalcone isomerase in plants
1
2022
... CHI 在花青素合成过程中也起着重要的催化作用,主要在查尔酮转化生成黄烷酮的过程中起催化作用.目前,已在番茄(Lycopersicon esculentum Mill.)[41 ] 、洋葱(Allium cepa )[56 ] 、大豆[Glycine max (Linn.) Merr.][57 ] 、菜豆(Phaseolus vulgaris L.)[58 ] 等植物中分离克隆出了CHI 基因,其表达对花青素以及类黄酮的积累有重要作用.将矮牵牛CHI 基因转入番茄中过度表达,转基因番茄果皮中类黄酮化合物含量增加78倍[41 ] .Morita等[59 ] 在日本牵牛花中鉴定出了一种花青素合成增强基因EFP ,发现敲除EFP 基因使牵牛花(Petunia hybrida )和斗牛花(Torenia hybrida )中花青素的含量显著降低,表现为花色褪去. ...
Overexpression of chalcone isomerase A gene in Astragalus trigonus for stimulating apigenin
1
2021
... CHI 在花青素合成过程中也起着重要的催化作用,主要在查尔酮转化生成黄烷酮的过程中起催化作用.目前,已在番茄(Lycopersicon esculentum Mill.)[41 ] 、洋葱(Allium cepa )[56 ] 、大豆[Glycine max (Linn.) Merr.][57 ] 、菜豆(Phaseolus vulgaris L.)[58 ] 等植物中分离克隆出了CHI 基因,其表达对花青素以及类黄酮的积累有重要作用.将矮牵牛CHI 基因转入番茄中过度表达,转基因番茄果皮中类黄酮化合物含量增加78倍[41 ] .Morita等[59 ] 在日本牵牛花中鉴定出了一种花青素合成增强基因EFP ,发现敲除EFP 基因使牵牛花(Petunia hybrida )和斗牛花(Torenia hybrida )中花青素的含量显著降低,表现为花色褪去. ...
A chalcone isomerase-like protein enhances flavonoid production and flower pigmentation
1
2014
... CHI 在花青素合成过程中也起着重要的催化作用,主要在查尔酮转化生成黄烷酮的过程中起催化作用.目前,已在番茄(Lycopersicon esculentum Mill.)[41 ] 、洋葱(Allium cepa )[56 ] 、大豆[Glycine max (Linn.) Merr.][57 ] 、菜豆(Phaseolus vulgaris L.)[58 ] 等植物中分离克隆出了CHI 基因,其表达对花青素以及类黄酮的积累有重要作用.将矮牵牛CHI 基因转入番茄中过度表达,转基因番茄果皮中类黄酮化合物含量增加78倍[41 ] .Morita等[59 ] 在日本牵牛花中鉴定出了一种花青素合成增强基因EFP ,发现敲除EFP 基因使牵牛花(Petunia hybrida )和斗牛花(Torenia hybrida )中花青素的含量显著降低,表现为花色褪去. ...
Tissue-specific accumulation and subcellular localization of chalcone isomerase (CHI) in grapevine
1
2019
... 有学者在研究过程中发现, CHI 主要存在于葡萄果实的表皮细胞的细胞质、细胞核及叶绿体中,而在葡萄果实的其他组织中,CHI 主要存在于细胞质、细胞壁和细胞核中,该基因存在表达特异性[60 ] . ...
Molecular characterization and expression of alfalfa (Medicago sativa L.) flavanone-3-hydroxylase and dihydroflavonol-4-reductase encoding genes
1
1995
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
‘红阳’猕猴桃cDNA文库构建及F3H基因的表达初探
1
2009
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
‘红阳’猕猴桃cDNA文库构建及F3H基因的表达初探
1
2009
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
Flavanone 3-hydroxylase transcripts and flavonol accumulation are temporally coordinate in maize anthers
1
1995
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
淹水胁迫对抗菌F3'H基因表达及其下游产物含量的影响
1
2018
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
淹水胁迫对抗菌F3'H基因表达及其下游产物含量的影响
1
2018
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
Cloning of the flavonoid 3'-hydroxylase gene of Eustoma grandiflorum (raf.) shinn. (EgF 3'H ) and complementation of an F3'H-deficient mutant of Ipomoea nil (L.) Roth. by heterologous expression of EgF 3'H
1
2015
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
Silencing of flavanone-3-hydroxylase in apple (Malus ×domestica Borkh.) leads to accumulation of flavanones, but not to reduced fire blight susceptibility
1
2012
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
RNAi-mediated silencing of the flavanone 3-hydroxylase gene and its effect on flavonoid biosynthesis in strawberry fruit
1
2013
... 黄烷酮-3-羟化酶属于一种加氧型酶,是调控花青素合成的重要酶基因,发挥作用时要依赖于Fe2+ 、氧等辅助因子.F3H是类黄酮等多个代谢合成途径中关键限速酶,黄烷酮经F 3H 催化转化为苯丙氨酸下游代谢物的直接前体物质——二氢黄酮醇.据1995年Charrier等[61 ] 报道了F 3H 基因序列以来,目前已在猕猴桃(Actinidia )[62 ] 、玉米(Zea mays L.)[63 ] 、菊花(Dendranthema morifolium )[64 ] 、桔梗(Platycodon grandiflorus )[65 ] 中克隆出了他们对应的F 3H 基因.在反义抑制黄烷酮-3-羟化酶的转基因苹果植株中,表现为阻断花青素以及类黄酮物质的合成[66 ] .Jiang等[67 ] 构建含有编码F 3H 基因的RNAi基因沉默载体后,将含有RNAi基因沉默载体的农杆菌导入授粉14 d的草莓10 d后,经检测与野生型相比F 3H 的相对表达量下调了70%,且花青素含量大幅度降低. ...
芒果二氢黄酮醇4-还原酶(DFR )基因的克隆及其表达分析
1
2015
... 无色的花色苷是二氢黄酮醇通过二氢黄酮醇-4-还原酶催化生成[68 ] .有研究表明,DFR 基因与CHI 基因相似,存在表达特异性.如凤兰(Cymbidium dayanum Rchb. F.)中,DFR 基因主要在幼叶中表达,而在花器官中几乎检测不到[69 ] .通过克隆百合(Lilium Asiatic )DFR酶编码基因LhDFR 并分析其时空表达,发现在白色鳞茎以及未着色的叶和茎中,LhDFR 基因均未表达,而在花药、花丝,以及雌蕊中大量表达[70 ] .DFR 的表达是一个复杂的调控网,调控方式需要进一步去探究. ...
芒果二氢黄酮醇4-还原酶(DFR )基因的克隆及其表达分析
1
2015
... 无色的花色苷是二氢黄酮醇通过二氢黄酮醇-4-还原酶催化生成[68 ] .有研究表明,DFR 基因与CHI 基因相似,存在表达特异性.如凤兰(Cymbidium dayanum Rchb. F.)中,DFR 基因主要在幼叶中表达,而在花器官中几乎检测不到[69 ] .通过克隆百合(Lilium Asiatic )DFR酶编码基因LhDFR 并分析其时空表达,发现在白色鳞茎以及未着色的叶和茎中,LhDFR 基因均未表达,而在花药、花丝,以及雌蕊中大量表达[70 ] .DFR 的表达是一个复杂的调控网,调控方式需要进一步去探究. ...
Cloning and characterization of full-length cDNA clones encoding chalcone synthase from the orchid Bromheadia finlaysoniana
1
1998
... 无色的花色苷是二氢黄酮醇通过二氢黄酮醇-4-还原酶催化生成[68 ] .有研究表明,DFR 基因与CHI 基因相似,存在表达特异性.如凤兰(Cymbidium dayanum Rchb. F.)中,DFR 基因主要在幼叶中表达,而在花器官中几乎检测不到[69 ] .通过克隆百合(Lilium Asiatic )DFR酶编码基因LhDFR 并分析其时空表达,发现在白色鳞茎以及未着色的叶和茎中,LhDFR 基因均未表达,而在花药、花丝,以及雌蕊中大量表达[70 ] .DFR 的表达是一个复杂的调控网,调控方式需要进一步去探究. ...
Spatial and temporal expression of chalcone synthase and dihydroflavonol 4-reductase genes in the Asiatic hybrid lily
1
2003
... 无色的花色苷是二氢黄酮醇通过二氢黄酮醇-4-还原酶催化生成[68 ] .有研究表明,DFR 基因与CHI 基因相似,存在表达特异性.如凤兰(Cymbidium dayanum Rchb. F.)中,DFR 基因主要在幼叶中表达,而在花器官中几乎检测不到[69 ] .通过克隆百合(Lilium Asiatic )DFR酶编码基因LhDFR 并分析其时空表达,发现在白色鳞茎以及未着色的叶和茎中,LhDFR 基因均未表达,而在花药、花丝,以及雌蕊中大量表达[70 ] .DFR 的表达是一个复杂的调控网,调控方式需要进一步去探究. ...
New insights into the regulation of anthocyanin biosynthesis in fruits
1
2013
... 花青苷合成酶是花青素生物合成过程中最后一步反应的关键酶,其作用主要将无色的花色素氧化生成有色但易分解的花色素[71 ] .对花青苷合成酶的研究发现其表达具有一定的品种或组织特异性.Li等[72 ] 测定了紫色和白色品种的桑果中ANS编码基因的表达量,发现ANS 基因只在紫色桑果果肉中高表达.在菠菜中,ANS 在大多数器官和组织中均未表达,且只在种子中检测到了ANS的活性[73 ] .利用RT-PCR技术从桃果皮中分离出了ANS编码基因PpANS ,对其在桃植株中的表达进行了研究,结果发现,PpANS 在桃花、果皮,以及果肉中高表达,其中以果皮中的表达量最高,而在桃植株的根、茎、叶中均未表达[74 ] .这些研究结果表明,花青素的合成与积累与ANS的表达有直接关系. ...
Isolation and expression analysis of anthocyanin biosynthetic genes in Morus alba L
2
2014
... 花青苷合成酶是花青素生物合成过程中最后一步反应的关键酶,其作用主要将无色的花色素氧化生成有色但易分解的花色素[71 ] .对花青苷合成酶的研究发现其表达具有一定的品种或组织特异性.Li等[72 ] 测定了紫色和白色品种的桑果中ANS编码基因的表达量,发现ANS 基因只在紫色桑果果肉中高表达.在菠菜中,ANS 在大多数器官和组织中均未表达,且只在种子中检测到了ANS的活性[73 ] .利用RT-PCR技术从桃果皮中分离出了ANS编码基因PpANS ,对其在桃植株中的表达进行了研究,结果发现,PpANS 在桃花、果皮,以及果肉中高表达,其中以果皮中的表达量最高,而在桃植株的根、茎、叶中均未表达[74 ] .这些研究结果表明,花青素的合成与积累与ANS的表达有直接关系. ...
... ABA作为一种植物激素参与了植物的多种生理过程,如种子休眠、气孔关闭以及光合作用等.此外, ABA还参与了跃变果实的成熟[115 ] ,而对非跃变果实的成熟作用不显著[116 ] .近年来,研究证明ABA不仅参与果实成熟,也参与调控果皮花青素的积累.Shen等[117 ] 用ABA的生物合成抑制剂去二氢愈创木酸(NDGA)和1 mmol·L-1 ABA处理樱桃,发现NDGA抑制了花青素的积累,ABA处理樱桃2 h后,检测到PacMYBA 的表达量达到了高峰,另外,构建一种烟草病毒(TRV)沉默樱桃ABA合成过程中的编码关键酶基因PacNCED 1,发现樱桃果实内源ABA的含量降低,以及花青素合成关键酶活性及其基因表达的下调,抑制了花青素的积累.对荔枝果实分别进行ABA和光照处理,研究结果表明两种处理均能促进LcMYB 1基因的表达,且在LcMYB 1启动子区鉴定了与光响应性和脱落酸响应性相关的顺式元件,数据显示,在花青素合成的结构基因中只有LcUFGT 与LcMYB 1高度相关[72 ] .目前,对于ABA调控花青素合成的机理还有待进一步研究. ...
Both HY5 and HYH are necessary regulators for low temperature-induced anthocyanin accumulation in Arabidopsis seedlings
1
2011
... 花青苷合成酶是花青素生物合成过程中最后一步反应的关键酶,其作用主要将无色的花色素氧化生成有色但易分解的花色素[71 ] .对花青苷合成酶的研究发现其表达具有一定的品种或组织特异性.Li等[72 ] 测定了紫色和白色品种的桑果中ANS编码基因的表达量,发现ANS 基因只在紫色桑果果肉中高表达.在菠菜中,ANS 在大多数器官和组织中均未表达,且只在种子中检测到了ANS的活性[73 ] .利用RT-PCR技术从桃果皮中分离出了ANS编码基因PpANS ,对其在桃植株中的表达进行了研究,结果发现,PpANS 在桃花、果皮,以及果肉中高表达,其中以果皮中的表达量最高,而在桃植株的根、茎、叶中均未表达[74 ] .这些研究结果表明,花青素的合成与积累与ANS的表达有直接关系. ...
Molecular cloning and characterization of an anthocyanidin synthase gene in Prunus persica (L.) batsch
2
2017
... 花青苷合成酶是花青素生物合成过程中最后一步反应的关键酶,其作用主要将无色的花色素氧化生成有色但易分解的花色素[71 ] .对花青苷合成酶的研究发现其表达具有一定的品种或组织特异性.Li等[72 ] 测定了紫色和白色品种的桑果中ANS编码基因的表达量,发现ANS 基因只在紫色桑果果肉中高表达.在菠菜中,ANS 在大多数器官和组织中均未表达,且只在种子中检测到了ANS的活性[73 ] .利用RT-PCR技术从桃果皮中分离出了ANS编码基因PpANS ,对其在桃植株中的表达进行了研究,结果发现,PpANS 在桃花、果皮,以及果肉中高表达,其中以果皮中的表达量最高,而在桃植株的根、茎、叶中均未表达[74 ] .这些研究结果表明,花青素的合成与积累与ANS的表达有直接关系. ...
... 外源ALA通过诱导花青素的积累,从而影响植物果实果皮花青素的含量.对葡萄喷施ALA,发现100 mg·L-1 的ALA显著增加了果皮花青素的含量同时也显著增加了PAL的活性,与对照水平相比,分别增加了194.71%、82.35%[112 ] .对苹果设置不同浓度梯度的ALA进行喷施,发现100 mg·L-1 ALA对苹果果皮花青素的积累量的影响最为显著,且在处理25 d后,与对照水平相比,苹果花青素合成仍表现出促进效应[113 ] .ALA对花青素的促进积累的效应还表现在对花青素生物合成关键酶基因的调控中.在桃表皮涂抹ALA,研究结果发现ALA不仅促进了桃果皮花青素含量,还显著提高了UFGT、DFR、LDOX 和 CHS等酶的活性,4个基因的表达量高峰也随之提前[114 ] ,在转录水平上,显著促进了桃果皮中MYB 10和WD 40基因的表达量,而BHLH3蛋白的相对表达量无显著影响,这些酶基因在转录水平与花青素的积累水平相一致[74 ] .对喷施ALA的离体葡萄果皮进行半定量RT-PCR,检测结果表明ALA能有效促进花青素合成过程中结构基因(PAL、CHS、CHI、DFR、UFGT)以及调控因子(MYB、bHLH3、WD40)的表达.而对于ALA对花青素合成的调控机制,可能是由于在果皮喷施ALA后,ALA的向下代谢成卟啉化合物来实现对花青素的调控,但具体的调控机制有待进一步研究. ...
Three R2R3-MYB transcription factors from banana (Musa acuminata ) activate structural anthocyanin biosynthesis genes as part of an MBW complex
1
2023
... 在花青素合成过程中,调控基因在转录水平上同样起到了决定性的作用.结构基因经转录因子调控以识别特异性结合的区域,影响花青素的生物合成[75 ] .转录因子MYB、bHLH和WD40主要调控花青素的合成.MYB 、bHLH 和WD 40基因在作用的过程中能够形成二元复合物或三元复合物,以此正向或负向调控花青素的合成[76 ] . ...
The apple WD40 protein MdTTG1 interacts with bHLH but not MYB proteins to regulate anthocyanin accumulation
2
2012
... 在花青素合成过程中,调控基因在转录水平上同样起到了决定性的作用.结构基因经转录因子调控以识别特异性结合的区域,影响花青素的生物合成[75 ] .转录因子MYB、bHLH和WD40主要调控花青素的合成.MYB 、bHLH 和WD 40基因在作用的过程中能够形成二元复合物或三元复合物,以此正向或负向调控花青素的合成[76 ] . ...
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators
1
1987
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
Characterization of the soybean R2R3-MYB transcription factor GmMYB 81 and its functional roles under abiotic stresses
1
2020
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
A MYB transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia mangostana L.) fruit during ripening
2
2009
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
An apple MYB transcription factor, MdMYB3, is involved in regulation of anthocyanin biosynthesis and flower development
1
2013
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
Effect of exogenous sucrose on anthocyanin synthesis in postharvest strawberry fruit
2
2019
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
A R2R3-MYB transcription factor, VvMYBC2L2, functions as a transcriptional repressor of anthocyanin biosynthesis in grapevine (Vitis vinifera L.)
1
2018
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
The atroviolacea gene encodes an R3-MYB protein repressing anthocyanin synthesis in tomato plants
1
2018
... 在高等植物中,MYB是极其重要的一类转录因子,参与了植物多种发育过程以及植物抗逆性的形成.1987年Paz-Ares等[77 ] 在玉米中分离并鉴定出了ZmC1,这是植物中被鉴定的第一个 MYB家族转录因子.后来,随着技术手段的成熟,相继在许多植物中鉴定出了MYB转录因子.目前,已在大豆中鉴定出了244种R2R3型MYBs,其中与两种非生物应激调节剂ATmyb 44和ATmyb 77密切相关的R2R3型MYBs(GmMYB 81)被证明在大豆组织和胚胎中差异表达,且在大豆逆境条件下的表达显著上调[78 ] .MYB转录因子家族各成员能够调控植物组织着色,在花青素合成的过程中发挥了重要的作用(表2 ).许多MYB型转录因子已经被证明参与了花青素生物合成的正向调控.在山竹(Garcinia mangostana L.)果皮中,研究者克隆得到GmMYB 1,GmMYB 7,GmMYB 10和GmUFGT ,发现GmMYB 10和GmUFGT 在山竹果实成熟和着色过程中高表达,促进了山竹花青素的积累[79 ] .在苹果中,MdMYB 3的过表达促进了CHS 、CHI 、UFGT 、FLS 等结构基因上调,且发现在红色果皮中MdMYB 3的表达高于黄色果皮苹果;另外,转MdMYB3的烟草植株比野生型的花颜色更深[80 ] .Li等[81 ] 在荔枝中(Litchi chinensis Sonn.)克隆并鉴定了一种新型R2R3-MYBS LcMYB5转录因子,发现在荔枝中的表达与其果皮花青素的积累无相关性关系,而在烟草和矮牵牛中过表达,能够通过直接激活花青素合成关键基因的表达促进其积累.另一方面,部分MYB转录因子也能够负向调控花青素的生物合成.在葡萄中发现了一个R2R3-MYB转录因子VvMYBC 2L 2能够负向调控花青素的生物合成.Zhu等[82 ] 发现VvMYBC2L2作为一种核蛋白,在葡萄果实发育过程中在果皮细胞中高表达,而果实到达转色期时表达降低,另外,花青素生物合成关键基因VvDFR 1和VvDLOX 1与VvMYBC 2L 2的表达模式相一致.Colanero等[83 ] 在番茄突变体atv 的候选基因中发现一种R3-MYB的突变基因,其过表达时,通过负调控内源MBW复合物来介导花青素合成,且可以直接和MBW中bHLH转录因子结合,形成底物竞争关系干扰番茄花青素的合成和积累. ...
A functional homologue of Arabidopsis TTG 1 from Freesia interacts with bHLH proteins to regulate anthocyanin and proanthocyanidin biosynthesis in both Freesia hybrida and Arabidopsis thaliana
1
2019
... bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana
1
2004
... bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The tomato hoffman’s anthocyaninless gene encodes a bHLH transcription factor involved in anthocyanin biosynthesis that is developmentally regulated and induced by low temperatures
2
2016
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
... bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
A MYB/bHLH complex regulates tissue-specific anthocyanin biosynthesis in the inner pericarp of red-centered kiwifruit Actinidia chinensis cv. Hongyang
3
2019
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
... bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
Dual DNA binding specificity of a petal epidermis-specific MYB transcription factor (MYB.Ph3) from Petunia hybrida
1
1995
... bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The Prp19 WD40 domain contains a conserved protein interaction region essential for its function
1
2010
... 转录因子在花青素合成过程中主要促进MYB-bHLH-WD40蛋白间的复合物的相互作用,从而为花青素的合成提供一个相对稳定的环境条件[89 ] .目前,有研究者已经从不同物种中分离并鉴定出WD40,另外,发现在多个物种中WD40基因家族中的AN11、TTG1、PAC1、PFWD等蛋白参与并调控了花青素合成整个过程的关键结构酶基因.De Vetten等[90 ] 分离了第一个植物体中的WD40蛋白——矮牵牛AN11,研究发现在矮牵牛an11突变体中,DFR 的表达量下调,另外在矮牵牛花瓣中,AN 2过表达后,AN 11恢复,上调了花青素结构基因的表达,促进了花青素的积累.在其后的研究中,在拟南芥中克隆并鉴定出了的矮牵牛AN 11同源基因TTG 1基因,两者具有高度的相似性[91 ] .在蓝莓中,VcMYBL 1、VcbHLH 1、VcWDL 2的表达与花青素的积累和颜色的变化呈正相关[92 ] .Meng等[93 ] 发现紫花苜蓿WHITE PETAL 1(WP 1)基因和MtTT 8基因通过和MtWD 40-1相互作用激活了类胡萝卜素生物合成基因MtLYCe 和MtLYCb ,进而促进了类胡萝卜素和花青素的合成.Kang等[94 ] 发现紫番茄中,果皮花青素的产生受MYB、bHLH和WD40转录因子的共同调节. ...
11 locus controlling flower pigmentation in petunia encodes a novel WD-repeat protein conserved in yeast, plants, and animals
2
1997
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
... 转录因子在花青素合成过程中主要促进MYB-bHLH-WD40蛋白间的复合物的相互作用,从而为花青素的合成提供一个相对稳定的环境条件[89 ] .目前,有研究者已经从不同物种中分离并鉴定出WD40,另外,发现在多个物种中WD40基因家族中的AN11、TTG1、PAC1、PFWD等蛋白参与并调控了花青素合成整个过程的关键结构酶基因.De Vetten等[90 ] 分离了第一个植物体中的WD40蛋白——矮牵牛AN11,研究发现在矮牵牛an11突变体中,DFR 的表达量下调,另外在矮牵牛花瓣中,AN 2过表达后,AN 11恢复,上调了花青素结构基因的表达,促进了花青素的积累.在其后的研究中,在拟南芥中克隆并鉴定出了的矮牵牛AN 11同源基因TTG 1基因,两者具有高度的相似性[91 ] .在蓝莓中,VcMYBL 1、VcbHLH 1、VcWDL 2的表达与花青素的积累和颜色的变化呈正相关[92 ] .Meng等[93 ] 发现紫花苜蓿WHITE PETAL 1(WP 1)基因和MtTT 8基因通过和MtWD 40-1相互作用激活了类胡萝卜素生物合成基因MtLYCe 和MtLYCb ,进而促进了类胡萝卜素和花青素的合成.Kang等[94 ] 发现紫番茄中,果皮花青素的产生受MYB、bHLH和WD40转录因子的共同调节. ...
Recent advances in the biosynthesis and accumulation of anthocyanins
1
2003
... 转录因子在花青素合成过程中主要促进MYB-bHLH-WD40蛋白间的复合物的相互作用,从而为花青素的合成提供一个相对稳定的环境条件[89 ] .目前,有研究者已经从不同物种中分离并鉴定出WD40,另外,发现在多个物种中WD40基因家族中的AN11、TTG1、PAC1、PFWD等蛋白参与并调控了花青素合成整个过程的关键结构酶基因.De Vetten等[90 ] 分离了第一个植物体中的WD40蛋白——矮牵牛AN11,研究发现在矮牵牛an11突变体中,DFR 的表达量下调,另外在矮牵牛花瓣中,AN 2过表达后,AN 11恢复,上调了花青素结构基因的表达,促进了花青素的积累.在其后的研究中,在拟南芥中克隆并鉴定出了的矮牵牛AN 11同源基因TTG 1基因,两者具有高度的相似性[91 ] .在蓝莓中,VcMYBL 1、VcbHLH 1、VcWDL 2的表达与花青素的积累和颜色的变化呈正相关[92 ] .Meng等[93 ] 发现紫花苜蓿WHITE PETAL 1(WP 1)基因和MtTT 8基因通过和MtWD 40-1相互作用激活了类胡萝卜素生物合成基因MtLYCe 和MtLYCb ,进而促进了类胡萝卜素和花青素的合成.Kang等[94 ] 发现紫番茄中,果皮花青素的产生受MYB、bHLH和WD40转录因子的共同调节. ...
Identification and characterization of MYB-bHLH-WD40 regulatory complex members controlling anthocyanidin biosynthesis in blueberry fruits development
1
2019
... 转录因子在花青素合成过程中主要促进MYB-bHLH-WD40蛋白间的复合物的相互作用,从而为花青素的合成提供一个相对稳定的环境条件[89 ] .目前,有研究者已经从不同物种中分离并鉴定出WD40,另外,发现在多个物种中WD40基因家族中的AN11、TTG1、PAC1、PFWD等蛋白参与并调控了花青素合成整个过程的关键结构酶基因.De Vetten等[90 ] 分离了第一个植物体中的WD40蛋白——矮牵牛AN11,研究发现在矮牵牛an11突变体中,DFR 的表达量下调,另外在矮牵牛花瓣中,AN 2过表达后,AN 11恢复,上调了花青素结构基因的表达,促进了花青素的积累.在其后的研究中,在拟南芥中克隆并鉴定出了的矮牵牛AN 11同源基因TTG 1基因,两者具有高度的相似性[91 ] .在蓝莓中,VcMYBL 1、VcbHLH 1、VcWDL 2的表达与花青素的积累和颜色的变化呈正相关[92 ] .Meng等[93 ] 发现紫花苜蓿WHITE PETAL 1(WP 1)基因和MtTT 8基因通过和MtWD 40-1相互作用激活了类胡萝卜素生物合成基因MtLYCe 和MtLYCb ,进而促进了类胡萝卜素和花青素的合成.Kang等[94 ] 发现紫番茄中,果皮花青素的产生受MYB、bHLH和WD40转录因子的共同调节. ...
The MYB activator WHITE PETAL1 associates with MtTT8 and MtWD40-1 to regulate carotenoid-derived flower pigmentation in Medicago truncatula
1
2019
... 转录因子在花青素合成过程中主要促进MYB-bHLH-WD40蛋白间的复合物的相互作用,从而为花青素的合成提供一个相对稳定的环境条件[89 ] .目前,有研究者已经从不同物种中分离并鉴定出WD40,另外,发现在多个物种中WD40基因家族中的AN11、TTG1、PAC1、PFWD等蛋白参与并调控了花青素合成整个过程的关键结构酶基因.De Vetten等[90 ] 分离了第一个植物体中的WD40蛋白——矮牵牛AN11,研究发现在矮牵牛an11突变体中,DFR 的表达量下调,另外在矮牵牛花瓣中,AN 2过表达后,AN 11恢复,上调了花青素结构基因的表达,促进了花青素的积累.在其后的研究中,在拟南芥中克隆并鉴定出了的矮牵牛AN 11同源基因TTG 1基因,两者具有高度的相似性[91 ] .在蓝莓中,VcMYBL 1、VcbHLH 1、VcWDL 2的表达与花青素的积累和颜色的变化呈正相关[92 ] .Meng等[93 ] 发现紫花苜蓿WHITE PETAL 1(WP 1)基因和MtTT 8基因通过和MtWD 40-1相互作用激活了类胡萝卜素生物合成基因MtLYCe 和MtLYCb ,进而促进了类胡萝卜素和花青素的合成.Kang等[94 ] 发现紫番茄中,果皮花青素的产生受MYB、bHLH和WD40转录因子的共同调节. ...
Expression of anthocyanin biosynthesis-related genes reflects the peel color in purple tomato
1
2018
... 转录因子在花青素合成过程中主要促进MYB-bHLH-WD40蛋白间的复合物的相互作用,从而为花青素的合成提供一个相对稳定的环境条件[89 ] .目前,有研究者已经从不同物种中分离并鉴定出WD40,另外,发现在多个物种中WD40基因家族中的AN11、TTG1、PAC1、PFWD等蛋白参与并调控了花青素合成整个过程的关键结构酶基因.De Vetten等[90 ] 分离了第一个植物体中的WD40蛋白——矮牵牛AN11,研究发现在矮牵牛an11突变体中,DFR 的表达量下调,另外在矮牵牛花瓣中,AN 2过表达后,AN 11恢复,上调了花青素结构基因的表达,促进了花青素的积累.在其后的研究中,在拟南芥中克隆并鉴定出了的矮牵牛AN 11同源基因TTG 1基因,两者具有高度的相似性[91 ] .在蓝莓中,VcMYBL 1、VcbHLH 1、VcWDL 2的表达与花青素的积累和颜色的变化呈正相关[92 ] .Meng等[93 ] 发现紫花苜蓿WHITE PETAL 1(WP 1)基因和MtTT 8基因通过和MtWD 40-1相互作用激活了类胡萝卜素生物合成基因MtLYCe 和MtLYCb ,进而促进了类胡萝卜素和花青素的合成.Kang等[94 ] 发现紫番茄中,果皮花青素的产生受MYB、bHLH和WD40转录因子的共同调节. ...
Isolation and functional characterization of a R2R3-MYB regulator of Prunus mume anthocyanin biosynthetic pathway
1
2017
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The onion (Allium cepa L.) R 2R 3-MYB gene MYB 1 regulates anthocyanin biosynthesis
1
2016
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The identification of a MYB transcription factor controlling anthocyanin biosynthesis regulation in Chrysanthemum flowers
1
2015
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
A wheat MYB transcriptional repressor TaMyb1D regulates phenylpropanoid metabolism and enhances tolerance to drought and oxidative stresses in transgenic tobacco plants
1
2017
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
A rice gene, OsPL , encoding a MYB family transcription factor confers anthocyanin synthesis, heat stress response and hormonal signaling
1
2019
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis
1
2008
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
A group of grapevine MYBA transcription factors located in chromosome 14 control anthocyanin synthesis in vegetative organs with different specificities compared with the berry color locus
1
2017
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
A MYB transcription factor, DcMYB6, is involved in regulating anthocyanin biosynthesis in purple carrot taproots
1
2017
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
Activator-type R2R3-MYB genes induce a repressor-type R2R3-MYB gene to balance anthocyanin and proanthocyanidin accumulation
2
2019
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
... [
103 ]
bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ] 2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
PsbHLH 1, a novel transcription factor involved in regulating anthocyanin biosynthesis in tree peony (Paeonia suffruticosa )
1
2020
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
A pomegranate (Punica granatum L.) WD40-repeat gene is a functional homologue of Arabidopsis TTG1 and is involved in the regulation of anthocyanin biosynthesis during pomegranate fruit development
2
2011
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
... [
105 ]
GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ] 2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
Regulation of anthocyanin biosynthesis by nitrogen in TTG1-GL3/TT8-PAP1-programmed red cells of Arabidopsis thaliana
1
2012
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The molecular cloning and functional characterization of MdMYC2 , a bHLH transcription factor in apple
1
2016
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
The bHLH transcription factor MdbHLH3 promotes anthocyanin accumulation and fruit colouration in response to low temperature in apples
1
2012
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
Brassica napus GLABRA3-1 promotes anthocyanin biosynthesis and trichome formation in true leaves when expressed in Arabidopsis thaliana
1
2018
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
Two WD-repeat genes from cotton are functional homologues of the Arabidopsis thaliana TRANSPARENT TESTA GLABRA1 (TTG 1) gene
1
2005
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
Plant Cell
1
2010
... Transcription factors related to anthocyanin synthesis pathway
Table 2 类别 转录因子 物种 调节基因 参考文献 MYB PmMYBa1 梅花Armeniaca mume Sieb. DFR , ANS [95 ] EsAN2 淫羊藿Epimedium brevicornu Maxim. CHIS , CHI , ANS [96 ] CmMYB6 菊花Dendranthema morifolium Tzvel. DFR [97 ] GmMYB10 山竹Garcinia mangostana L. DFR , UFGT [79 ] TaMyb1D 烟草Nicotiana tabacum L. PAL , CHS , CHI , F 3H , DFR , FLS [98 ] OsPL 水稻Oryza sativa L. PAL , CHS , ANS [99 ] AtMYBL2 拟南芥Arabidopsis thaliana AtTT 8, DFR [100 ] TaPL1 小麦Triticum aestivum PAL , CHS , CHI , F 3H , DFR [37 ] VvMYBA1/6/7 葡萄Vitis vinifera L. UFGT , 3AT [101 ] DcMYB6 黑胡萝卜Daucus carota L. CHS , DFR [102 ] PpMYB10.1 桃Prunus persica DFR , UFGT [103 ] PpMYBPA1 桃Prunus persica LAR 1[103 ] bHLH PsbHLH1 牡丹Paeonia suffruticosa Andr. DFR , ANS [104 ] PubHLH2 石榴Punica granatum L. DFR , ANS [105 ] AtGL3/TT8 拟南芥Arabidopsis thaliana CHS , CHI , F 3H , F 3'H , DFR , ANS , GT [106 ] LeAH 番茄Lycopersicon esculentum Mill. F 3'5'H , DFR , ANS , 3GT , GST [86 ] MdMYC2 苹果Malus ×domestica DFR , UF 3GT , F 3H , CHS [107 ] MdbHLH3 苹果Malus ×domestica DFR , UFGT [108 ] AcbHLH42 猕猴桃Actinidia chinensis Planch F 3GT 1, ANS [87 ] SmTT8 茄子Solanum melongena L. CHI , F 3H , DFR , 3GT , 5GT [109 ] WD40 MdTTG1 苹果Malus ×domestica PAL , CHI , CHS [76 ] PhAN11 矮牵牛Petunia hybrida Vilm. DFR [90 ] Pu TTG1 石榴Punica granatum L. DFR , LDOX [105 ] GhTTG1/TTG3 棉花Gossypium spp. DFR [110 ] StAN11 马铃薯Solanum tuberosum DFR [111 ]
2.1.2 bHLH bHLH也是植物中重要的一类转录因子,其结构存在两个高度保守且功能不同的功能区,一个是C端的螺旋-环-螺旋(bHLH),另一个为N端的碱性区域.目前,大部分关于bHLH的研究都关注于其对植物抗性的效果以及参与信号转导的作用,对花青素的生物合成影响也是其中一个重要的部分.Shan等[84 ] 通过克隆小苍兰(Freesia hybrida )WD40基因家族的调节蛋白FhTTG 1并分析其功能,结果表明FhTTG 1与bHLH 两个非等位基因相互作用显著激活调控花青素合成相关的启动子,进而促进花青素或原花青素的积累.在拟南芥种皮中,编码原花青素的酶基因TT 2、TT 8、TTG 1与MBW复合体互作,bHLH调控TTG 1发挥作用[85 ] .在缺失bHLH 蛋白的番茄中,对应bHLH 蛋白位点的ahin FMTT 271基因研究发现,ahin FMTT 271的过表达上调了花青素生物合成关键酶基因,提高了花青素在幼苗下胚轴、叶片和果皮中的积累水平[86 ] .Wang等[87 ] 发现猕猴桃AcMYB 123和AcbHLH 42的组合作用,激活了编码花青素合成AcANS和AcF3GT1的启动子,且通过对烟草和拟南芥瞬时表达分析表明,AcMYB 123和AcbHLH 42的共同表达是花青素生物合成的必要条件.bHLH对花青素生物合成的调控是一个复杂的网络,但有趣的是,并不是所有植物的花青素合成都需要bHLH的参与,早在1995年Solano等[88 ] 在对矮牵牛的研究中发现,MYB3可以激活CHS的表达,但在作用的过程中不需要bHLH的协作. ...
外源ALA对克瑞森无核葡萄叶片光合特性及果实品质的影响
1
2018
... 外源ALA通过诱导花青素的积累,从而影响植物果实果皮花青素的含量.对葡萄喷施ALA,发现100 mg·L-1 的ALA显著增加了果皮花青素的含量同时也显著增加了PAL的活性,与对照水平相比,分别增加了194.71%、82.35%[112 ] .对苹果设置不同浓度梯度的ALA进行喷施,发现100 mg·L-1 ALA对苹果果皮花青素的积累量的影响最为显著,且在处理25 d后,与对照水平相比,苹果花青素合成仍表现出促进效应[113 ] .ALA对花青素的促进积累的效应还表现在对花青素生物合成关键酶基因的调控中.在桃表皮涂抹ALA,研究结果发现ALA不仅促进了桃果皮花青素含量,还显著提高了UFGT、DFR、LDOX 和 CHS等酶的活性,4个基因的表达量高峰也随之提前[114 ] ,在转录水平上,显著促进了桃果皮中MYB 10和WD 40基因的表达量,而BHLH3蛋白的相对表达量无显著影响,这些酶基因在转录水平与花青素的积累水平相一致[74 ] .对喷施ALA的离体葡萄果皮进行半定量RT-PCR,检测结果表明ALA能有效促进花青素合成过程中结构基因(PAL、CHS、CHI、DFR、UFGT)以及调控因子(MYB、bHLH3、WD40)的表达.而对于ALA对花青素合成的调控机制,可能是由于在果皮喷施ALA后,ALA的向下代谢成卟啉化合物来实现对花青素的调控,但具体的调控机制有待进一步研究. ...
外源ALA对克瑞森无核葡萄叶片光合特性及果实品质的影响
1
2018
... 外源ALA通过诱导花青素的积累,从而影响植物果实果皮花青素的含量.对葡萄喷施ALA,发现100 mg·L-1 的ALA显著增加了果皮花青素的含量同时也显著增加了PAL的活性,与对照水平相比,分别增加了194.71%、82.35%[112 ] .对苹果设置不同浓度梯度的ALA进行喷施,发现100 mg·L-1 ALA对苹果果皮花青素的积累量的影响最为显著,且在处理25 d后,与对照水平相比,苹果花青素合成仍表现出促进效应[113 ] .ALA对花青素的促进积累的效应还表现在对花青素生物合成关键酶基因的调控中.在桃表皮涂抹ALA,研究结果发现ALA不仅促进了桃果皮花青素含量,还显著提高了UFGT、DFR、LDOX 和 CHS等酶的活性,4个基因的表达量高峰也随之提前[114 ] ,在转录水平上,显著促进了桃果皮中MYB 10和WD 40基因的表达量,而BHLH3蛋白的相对表达量无显著影响,这些酶基因在转录水平与花青素的积累水平相一致[74 ] .对喷施ALA的离体葡萄果皮进行半定量RT-PCR,检测结果表明ALA能有效促进花青素合成过程中结构基因(PAL、CHS、CHI、DFR、UFGT)以及调控因子(MYB、bHLH3、WD40)的表达.而对于ALA对花青素合成的调控机制,可能是由于在果皮喷施ALA后,ALA的向下代谢成卟啉化合物来实现对花青素的调控,但具体的调控机制有待进一步研究. ...
Proteomics and SSH analyses of ALA-promoted fruit coloration and evidence for the involvement of a MADS-box gene, MdMADS 1
1
2016
... 外源ALA通过诱导花青素的积累,从而影响植物果实果皮花青素的含量.对葡萄喷施ALA,发现100 mg·L-1 的ALA显著增加了果皮花青素的含量同时也显著增加了PAL的活性,与对照水平相比,分别增加了194.71%、82.35%[112 ] .对苹果设置不同浓度梯度的ALA进行喷施,发现100 mg·L-1 ALA对苹果果皮花青素的积累量的影响最为显著,且在处理25 d后,与对照水平相比,苹果花青素合成仍表现出促进效应[113 ] .ALA对花青素的促进积累的效应还表现在对花青素生物合成关键酶基因的调控中.在桃表皮涂抹ALA,研究结果发现ALA不仅促进了桃果皮花青素含量,还显著提高了UFGT、DFR、LDOX 和 CHS等酶的活性,4个基因的表达量高峰也随之提前[114 ] ,在转录水平上,显著促进了桃果皮中MYB 10和WD 40基因的表达量,而BHLH3蛋白的相对表达量无显著影响,这些酶基因在转录水平与花青素的积累水平相一致[74 ] .对喷施ALA的离体葡萄果皮进行半定量RT-PCR,检测结果表明ALA能有效促进花青素合成过程中结构基因(PAL、CHS、CHI、DFR、UFGT)以及调控因子(MYB、bHLH3、WD40)的表达.而对于ALA对花青素合成的调控机制,可能是由于在果皮喷施ALA后,ALA的向下代谢成卟啉化合物来实现对花青素的调控,但具体的调控机制有待进一步研究. ...
5-氨基乙酰丙酸可促进桃果皮提前着色
1
2013
... 外源ALA通过诱导花青素的积累,从而影响植物果实果皮花青素的含量.对葡萄喷施ALA,发现100 mg·L-1 的ALA显著增加了果皮花青素的含量同时也显著增加了PAL的活性,与对照水平相比,分别增加了194.71%、82.35%[112 ] .对苹果设置不同浓度梯度的ALA进行喷施,发现100 mg·L-1 ALA对苹果果皮花青素的积累量的影响最为显著,且在处理25 d后,与对照水平相比,苹果花青素合成仍表现出促进效应[113 ] .ALA对花青素的促进积累的效应还表现在对花青素生物合成关键酶基因的调控中.在桃表皮涂抹ALA,研究结果发现ALA不仅促进了桃果皮花青素含量,还显著提高了UFGT、DFR、LDOX 和 CHS等酶的活性,4个基因的表达量高峰也随之提前[114 ] ,在转录水平上,显著促进了桃果皮中MYB 10和WD 40基因的表达量,而BHLH3蛋白的相对表达量无显著影响,这些酶基因在转录水平与花青素的积累水平相一致[74 ] .对喷施ALA的离体葡萄果皮进行半定量RT-PCR,检测结果表明ALA能有效促进花青素合成过程中结构基因(PAL、CHS、CHI、DFR、UFGT)以及调控因子(MYB、bHLH3、WD40)的表达.而对于ALA对花青素合成的调控机制,可能是由于在果皮喷施ALA后,ALA的向下代谢成卟啉化合物来实现对花青素的调控,但具体的调控机制有待进一步研究. ...
5-氨基乙酰丙酸可促进桃果皮提前着色
1
2013
... 外源ALA通过诱导花青素的积累,从而影响植物果实果皮花青素的含量.对葡萄喷施ALA,发现100 mg·L-1 的ALA显著增加了果皮花青素的含量同时也显著增加了PAL的活性,与对照水平相比,分别增加了194.71%、82.35%[112 ] .对苹果设置不同浓度梯度的ALA进行喷施,发现100 mg·L-1 ALA对苹果果皮花青素的积累量的影响最为显著,且在处理25 d后,与对照水平相比,苹果花青素合成仍表现出促进效应[113 ] .ALA对花青素的促进积累的效应还表现在对花青素生物合成关键酶基因的调控中.在桃表皮涂抹ALA,研究结果发现ALA不仅促进了桃果皮花青素含量,还显著提高了UFGT、DFR、LDOX 和 CHS等酶的活性,4个基因的表达量高峰也随之提前[114 ] ,在转录水平上,显著促进了桃果皮中MYB 10和WD 40基因的表达量,而BHLH3蛋白的相对表达量无显著影响,这些酶基因在转录水平与花青素的积累水平相一致[74 ] .对喷施ALA的离体葡萄果皮进行半定量RT-PCR,检测结果表明ALA能有效促进花青素合成过程中结构基因(PAL、CHS、CHI、DFR、UFGT)以及调控因子(MYB、bHLH3、WD40)的表达.而对于ALA对花青素合成的调控机制,可能是由于在果皮喷施ALA后,ALA的向下代谢成卟啉化合物来实现对花青素的调控,但具体的调控机制有待进一步研究. ...
Crop load and plant water status influence the ripening rate and aroma development in berries of grapevine (Vitis vinifera L.) cv. cabernet sauvignon
1
2021
... ABA作为一种植物激素参与了植物的多种生理过程,如种子休眠、气孔关闭以及光合作用等.此外, ABA还参与了跃变果实的成熟[115 ] ,而对非跃变果实的成熟作用不显著[116 ] .近年来,研究证明ABA不仅参与果实成熟,也参与调控果皮花青素的积累.Shen等[117 ] 用ABA的生物合成抑制剂去二氢愈创木酸(NDGA)和1 mmol·L-1 ABA处理樱桃,发现NDGA抑制了花青素的积累,ABA处理樱桃2 h后,检测到PacMYBA 的表达量达到了高峰,另外,构建一种烟草病毒(TRV)沉默樱桃ABA合成过程中的编码关键酶基因PacNCED 1,发现樱桃果实内源ABA的含量降低,以及花青素合成关键酶活性及其基因表达的下调,抑制了花青素的积累.对荔枝果实分别进行ABA和光照处理,研究结果表明两种处理均能促进LcMYB 1基因的表达,且在LcMYB 1启动子区鉴定了与光响应性和脱落酸响应性相关的顺式元件,数据显示,在花青素合成的结构基因中只有LcUFGT 与LcMYB 1高度相关[72 ] .目前,对于ABA调控花青素合成的机理还有待进一步研究. ...
Involvement of abscisic acid in postharvest water-deficit stress associated with the accumulation of anthocyanins in strawberry fruit
1
2016
... ABA作为一种植物激素参与了植物的多种生理过程,如种子休眠、气孔关闭以及光合作用等.此外, ABA还参与了跃变果实的成熟[115 ] ,而对非跃变果实的成熟作用不显著[116 ] .近年来,研究证明ABA不仅参与果实成熟,也参与调控果皮花青素的积累.Shen等[117 ] 用ABA的生物合成抑制剂去二氢愈创木酸(NDGA)和1 mmol·L-1 ABA处理樱桃,发现NDGA抑制了花青素的积累,ABA处理樱桃2 h后,检测到PacMYBA 的表达量达到了高峰,另外,构建一种烟草病毒(TRV)沉默樱桃ABA合成过程中的编码关键酶基因PacNCED 1,发现樱桃果实内源ABA的含量降低,以及花青素合成关键酶活性及其基因表达的下调,抑制了花青素的积累.对荔枝果实分别进行ABA和光照处理,研究结果表明两种处理均能促进LcMYB 1基因的表达,且在LcMYB 1启动子区鉴定了与光响应性和脱落酸响应性相关的顺式元件,数据显示,在花青素合成的结构基因中只有LcUFGT 与LcMYB 1高度相关[72 ] .目前,对于ABA调控花青素合成的机理还有待进一步研究. ...
A role for PacMYBA in ABA-regulated anthocyanin biosynthesis in red-colored sweet cherry cv. Hong Deng (Prunus avium L.)
1
2014
... ABA作为一种植物激素参与了植物的多种生理过程,如种子休眠、气孔关闭以及光合作用等.此外, ABA还参与了跃变果实的成熟[115 ] ,而对非跃变果实的成熟作用不显著[116 ] .近年来,研究证明ABA不仅参与果实成熟,也参与调控果皮花青素的积累.Shen等[117 ] 用ABA的生物合成抑制剂去二氢愈创木酸(NDGA)和1 mmol·L-1 ABA处理樱桃,发现NDGA抑制了花青素的积累,ABA处理樱桃2 h后,检测到PacMYBA 的表达量达到了高峰,另外,构建一种烟草病毒(TRV)沉默樱桃ABA合成过程中的编码关键酶基因PacNCED 1,发现樱桃果实内源ABA的含量降低,以及花青素合成关键酶活性及其基因表达的下调,抑制了花青素的积累.对荔枝果实分别进行ABA和光照处理,研究结果表明两种处理均能促进LcMYB 1基因的表达,且在LcMYB 1启动子区鉴定了与光响应性和脱落酸响应性相关的顺式元件,数据显示,在花青素合成的结构基因中只有LcUFGT 与LcMYB 1高度相关[72 ] .目前,对于ABA调控花青素合成的机理还有待进一步研究. ...
拟南芥转录因子Ethylene-insensitive3(EIN3 )抑制花青素的合成
1
2018
... 植物的成熟和衰老过程都有乙烯的参与以及调控.在乙烯信号转导的过程中,EIN3和EIL1两个蛋白发挥着重要的转录调控作用.有研究表明敲除拟南芥中ein 3和eil 1基因后,双突变体拟南芥的种子和叶片均表现为紫色,与单突变体ein3-1和野生型拟南芥相比较,其花青素含量显著增加[118 ] .同时,在野生型拟南芥体内乙烯抑制糖信号诱导的花青素的合成过程中,发现糖信号被抑制的同时抑制了其相关转录因子MYBL 2的表达,与此同时,一类影响花青素合成的负调控因子R3-MYB表达量提高,从而抑制了花青素的积累[119 ] .El-Kereamy等[120 ] 在葡萄表皮喷施乙烯释放物2-CEPA,24 h后葡萄内源乙烯含量与对照相比较增加了6倍,与此同时,2-CEPA还上调了花青素合成关键酶CHI和F3H的转录水平.乙烯在参与果实成熟过程中调控了果实花青素的合成,但果实中乙烯是直接作用于关键基因还是间接刺激转录因子来调控花青素的积累,具体的调控机制是一项亟待探究的课题. ...
拟南芥转录因子Ethylene-insensitive3(EIN3 )抑制花青素的合成
1
2018
... 植物的成熟和衰老过程都有乙烯的参与以及调控.在乙烯信号转导的过程中,EIN3和EIL1两个蛋白发挥着重要的转录调控作用.有研究表明敲除拟南芥中ein 3和eil 1基因后,双突变体拟南芥的种子和叶片均表现为紫色,与单突变体ein3-1和野生型拟南芥相比较,其花青素含量显著增加[118 ] .同时,在野生型拟南芥体内乙烯抑制糖信号诱导的花青素的合成过程中,发现糖信号被抑制的同时抑制了其相关转录因子MYBL 2的表达,与此同时,一类影响花青素合成的负调控因子R3-MYB表达量提高,从而抑制了花青素的积累[119 ] .El-Kereamy等[120 ] 在葡萄表皮喷施乙烯释放物2-CEPA,24 h后葡萄内源乙烯含量与对照相比较增加了6倍,与此同时,2-CEPA还上调了花青素合成关键酶CHI和F3H的转录水平.乙烯在参与果实成熟过程中调控了果实花青素的合成,但果实中乙烯是直接作用于关键基因还是间接刺激转录因子来调控花青素的积累,具体的调控机制是一项亟待探究的课题. ...
Ethylene suppression of sugar-induced anthocyanin pigmentation in Arabidopsis
1
2010
... 植物的成熟和衰老过程都有乙烯的参与以及调控.在乙烯信号转导的过程中,EIN3和EIL1两个蛋白发挥着重要的转录调控作用.有研究表明敲除拟南芥中ein 3和eil 1基因后,双突变体拟南芥的种子和叶片均表现为紫色,与单突变体ein3-1和野生型拟南芥相比较,其花青素含量显著增加[118 ] .同时,在野生型拟南芥体内乙烯抑制糖信号诱导的花青素的合成过程中,发现糖信号被抑制的同时抑制了其相关转录因子MYBL 2的表达,与此同时,一类影响花青素合成的负调控因子R3-MYB表达量提高,从而抑制了花青素的积累[119 ] .El-Kereamy等[120 ] 在葡萄表皮喷施乙烯释放物2-CEPA,24 h后葡萄内源乙烯含量与对照相比较增加了6倍,与此同时,2-CEPA还上调了花青素合成关键酶CHI和F3H的转录水平.乙烯在参与果实成熟过程中调控了果实花青素的合成,但果实中乙烯是直接作用于关键基因还是间接刺激转录因子来调控花青素的积累,具体的调控机制是一项亟待探究的课题. ...
Exogenous ethylene stimulates the long-term expression of genes related to anthocyanin biosynthesis in grape berries
1
2003
... 植物的成熟和衰老过程都有乙烯的参与以及调控.在乙烯信号转导的过程中,EIN3和EIL1两个蛋白发挥着重要的转录调控作用.有研究表明敲除拟南芥中ein 3和eil 1基因后,双突变体拟南芥的种子和叶片均表现为紫色,与单突变体ein3-1和野生型拟南芥相比较,其花青素含量显著增加[118 ] .同时,在野生型拟南芥体内乙烯抑制糖信号诱导的花青素的合成过程中,发现糖信号被抑制的同时抑制了其相关转录因子MYBL 2的表达,与此同时,一类影响花青素合成的负调控因子R3-MYB表达量提高,从而抑制了花青素的积累[119 ] .El-Kereamy等[120 ] 在葡萄表皮喷施乙烯释放物2-CEPA,24 h后葡萄内源乙烯含量与对照相比较增加了6倍,与此同时,2-CEPA还上调了花青素合成关键酶CHI和F3H的转录水平.乙烯在参与果实成熟过程中调控了果实花青素的合成,但果实中乙烯是直接作用于关键基因还是间接刺激转录因子来调控花青素的积累,具体的调控机制是一项亟待探究的课题. ...
Chemiluminometric evaluation of melatonin and selected melatonin precursors’ interaction with reactive oxygen and nitrogen species
1
2012
... 褪黑素作为色氨酸的一种衍生物类激素,近年来在农业生产中得到了广泛的运用[121 ] .有学者发现,在寒冷条件下,野生型拟南芥与缺少褪黑素生物合成酶基因的拟南芥突变体snat 的叶片的颜色更深,这一发现揭示了褪黑素能正向促进花青素的积累[122 ] .将草莓果实分别在50 mol·L-1 和100 mol·L-1 的褪黑素浸泡30 min后,测定出其花青素合成相关调控基因表达量上调,果实花青素积累增多,说明外源褪黑素可以正向调控苯丙氨酸代谢过程[123 ] .在Wei等[124 ] 对海棠的研究中表明,外源喷施褪黑素可以上调相关合成酶基因和转录因子的表达量,进而提高花青素的含量.有趣的是,有研究发现,充足的ATP和PEP可以有效增加甘薯和双孢菇PAL的活性,且可以增加花青素的积累量[125 -126 ] .有研究者推测,外源褪黑素可能促进了TCA循环中柠檬酸合成酶活性并且上调了柠檬酸合成相关基因的表达,为花青素合成提供更为丰富的合成底物,以此促进花青素的生物合成,但具体的调控机制仍为一项研究薄弱点. ...
Melatonin improved anthocyanin accumulation by regulating gene expressions and resulted in high reactive oxygen species scavenging capacity in cabbage
1
2016
... 褪黑素作为色氨酸的一种衍生物类激素,近年来在农业生产中得到了广泛的运用[121 ] .有学者发现,在寒冷条件下,野生型拟南芥与缺少褪黑素生物合成酶基因的拟南芥突变体snat 的叶片的颜色更深,这一发现揭示了褪黑素能正向促进花青素的积累[122 ] .将草莓果实分别在50 mol·L-1 和100 mol·L-1 的褪黑素浸泡30 min后,测定出其花青素合成相关调控基因表达量上调,果实花青素积累增多,说明外源褪黑素可以正向调控苯丙氨酸代谢过程[123 ] .在Wei等[124 ] 对海棠的研究中表明,外源喷施褪黑素可以上调相关合成酶基因和转录因子的表达量,进而提高花青素的含量.有趣的是,有研究发现,充足的ATP和PEP可以有效增加甘薯和双孢菇PAL的活性,且可以增加花青素的积累量[125 -126 ] .有研究者推测,外源褪黑素可能促进了TCA循环中柠檬酸合成酶活性并且上调了柠檬酸合成相关基因的表达,为花青素合成提供更为丰富的合成底物,以此促进花青素的生物合成,但具体的调控机制仍为一项研究薄弱点. ...
Insights into exogenous melatonin associated with phenylalanine metabolism in postharvest strawberry
1
2020
... 褪黑素作为色氨酸的一种衍生物类激素,近年来在农业生产中得到了广泛的运用[121 ] .有学者发现,在寒冷条件下,野生型拟南芥与缺少褪黑素生物合成酶基因的拟南芥突变体snat 的叶片的颜色更深,这一发现揭示了褪黑素能正向促进花青素的积累[122 ] .将草莓果实分别在50 mol·L-1 和100 mol·L-1 的褪黑素浸泡30 min后,测定出其花青素合成相关调控基因表达量上调,果实花青素积累增多,说明外源褪黑素可以正向调控苯丙氨酸代谢过程[123 ] .在Wei等[124 ] 对海棠的研究中表明,外源喷施褪黑素可以上调相关合成酶基因和转录因子的表达量,进而提高花青素的含量.有趣的是,有研究发现,充足的ATP和PEP可以有效增加甘薯和双孢菇PAL的活性,且可以增加花青素的积累量[125 -126 ] .有研究者推测,外源褪黑素可能促进了TCA循环中柠檬酸合成酶活性并且上调了柠檬酸合成相关基因的表达,为花青素合成提供更为丰富的合成底物,以此促进花青素的生物合成,但具体的调控机制仍为一项研究薄弱点. ...
Melatonin increases the performance of Malus hupehensis after UV-B exposure
1
2019
... 褪黑素作为色氨酸的一种衍生物类激素,近年来在农业生产中得到了广泛的运用[121 ] .有学者发现,在寒冷条件下,野生型拟南芥与缺少褪黑素生物合成酶基因的拟南芥突变体snat 的叶片的颜色更深,这一发现揭示了褪黑素能正向促进花青素的积累[122 ] .将草莓果实分别在50 mol·L-1 和100 mol·L-1 的褪黑素浸泡30 min后,测定出其花青素合成相关调控基因表达量上调,果实花青素积累增多,说明外源褪黑素可以正向调控苯丙氨酸代谢过程[123 ] .在Wei等[124 ] 对海棠的研究中表明,外源喷施褪黑素可以上调相关合成酶基因和转录因子的表达量,进而提高花青素的含量.有趣的是,有研究发现,充足的ATP和PEP可以有效增加甘薯和双孢菇PAL的活性,且可以增加花青素的积累量[125 -126 ] .有研究者推测,外源褪黑素可能促进了TCA循环中柠檬酸合成酶活性并且上调了柠檬酸合成相关基因的表达,为花青素合成提供更为丰富的合成底物,以此促进花青素的生物合成,但具体的调控机制仍为一项研究薄弱点. ...
Proteomic approach reveals that starch degradation contributes to anthocyanin accumulation in tuberous root of purple sweet potato
1
2016
... 褪黑素作为色氨酸的一种衍生物类激素,近年来在农业生产中得到了广泛的运用[121 ] .有学者发现,在寒冷条件下,野生型拟南芥与缺少褪黑素生物合成酶基因的拟南芥突变体snat 的叶片的颜色更深,这一发现揭示了褪黑素能正向促进花青素的积累[122 ] .将草莓果实分别在50 mol·L-1 和100 mol·L-1 的褪黑素浸泡30 min后,测定出其花青素合成相关调控基因表达量上调,果实花青素积累增多,说明外源褪黑素可以正向调控苯丙氨酸代谢过程[123 ] .在Wei等[124 ] 对海棠的研究中表明,外源喷施褪黑素可以上调相关合成酶基因和转录因子的表达量,进而提高花青素的含量.有趣的是,有研究发现,充足的ATP和PEP可以有效增加甘薯和双孢菇PAL的活性,且可以增加花青素的积累量[125 -126 ] .有研究者推测,外源褪黑素可能促进了TCA循环中柠檬酸合成酶活性并且上调了柠檬酸合成相关基因的表达,为花青素合成提供更为丰富的合成底物,以此促进花青素的生物合成,但具体的调控机制仍为一项研究薄弱点. ...
Exogenous adenosine triphosphate application retards cap browning in Agaricus bisporus during low temperature storage
1
2019
... 褪黑素作为色氨酸的一种衍生物类激素,近年来在农业生产中得到了广泛的运用[121 ] .有学者发现,在寒冷条件下,野生型拟南芥与缺少褪黑素生物合成酶基因的拟南芥突变体snat 的叶片的颜色更深,这一发现揭示了褪黑素能正向促进花青素的积累[122 ] .将草莓果实分别在50 mol·L-1 和100 mol·L-1 的褪黑素浸泡30 min后,测定出其花青素合成相关调控基因表达量上调,果实花青素积累增多,说明外源褪黑素可以正向调控苯丙氨酸代谢过程[123 ] .在Wei等[124 ] 对海棠的研究中表明,外源喷施褪黑素可以上调相关合成酶基因和转录因子的表达量,进而提高花青素的含量.有趣的是,有研究发现,充足的ATP和PEP可以有效增加甘薯和双孢菇PAL的活性,且可以增加花青素的积累量[125 -126 ] .有研究者推测,外源褪黑素可能促进了TCA循环中柠檬酸合成酶活性并且上调了柠檬酸合成相关基因的表达,为花青素合成提供更为丰富的合成底物,以此促进花青素的生物合成,但具体的调控机制仍为一项研究薄弱点. ...
Sugar sensing and signalling networks in plants
1
2005
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
The effect of ripening rates on the composition of Cabernet Sauvignon and Riesling wines: further insights into the sugar/flavor nexus
1
2022
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
苹果蔗糖转运蛋白MdSUT2调控花青苷积累的研究
1
2019
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
苹果蔗糖转运蛋白MdSUT2调控花青苷积累的研究
1
2019
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
光照对非光敏型茄子花青素合成相关基因的影响
1
2017
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
光照对非光敏型茄子花青素合成相关基因的影响
1
2017
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
5-氨基乙酰丙酸和金雀异黄素促进苹果果皮花青素形成的效应
1
2006
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
5-氨基乙酰丙酸和金雀异黄素促进苹果果皮花青素形成的效应
1
2006
... 糖作为一种信号转导物质已在多个领域研究和应用.对植物的生长和发育过程也起重要作用[127 ] .糖参与了植物的多种代谢过程,调控花青素生物合成过程中的大部分结构基因和调控的表达,如PAL、CHS、DFR、UFHT等[128 ] .Li等[81 ] 将草莓用50 mmol·L-1 的蔗糖溶液处理后,发现草莓果实果肉中花青素含量增加;Wang等[87 ] 发现,在光照条件下拟南芥经蔗糖溶液处理后,能显著诱导拟南芥花青素的积累.另外,植物内源蔗糖转运载体也能够参与花青素合成和积累途径.利用农杆菌转化法将在苹果中克隆的蔗糖转运载体基因MdSUT 2转化到拟南芥中,发现MdSUT 2的过表达会显著增加拟南芥花青素含量,而抑制MdSUT 2的表达则会降低花青素含量,说明蔗糖转运蛋白参与并调控花青素的生物合成[129 ] .在苹果中,MdSnRK 1.1与茉莉酸信号途径中的抑制因子MdJAZ 18相互作用,磷酸化后促进了26S蛋白酶介导的降解,调控了花青素生物合成相关基因的表达,并促进了花青素的积累[130 ] .MdJAZ 18、MdMYB 1、MdMYB 9和MdMYB 11均能与MdbHLH 3相互作用,MdbHLH 3不仅能上调MdJAZ 18、MdMYB 9和MdMYB 11的表达,并且激活了花青素生物合成基因MdDFR 、MdUFGT 、MdANS 、MdANR 和MdLAR 和调控基因MdMYB 1的表达,促进了原花青素和花青素的积累[17 ,131 ] .葡萄糖是植物光合作用的主要产物,同时也是参与细胞内众多生理生化过程的重要能源物质和碳源,而作为葡萄糖信号转导物质,苹果己糖激酶MdHXK1也参与了葡萄糖的感应和花青素的调控,其在调控的过程也是通过激活MBW复合蛋白和靶基因的表达来完成. ...
The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress
1
2015
... 光照是非生物环境因子中影响花青素生物合成的关键因子,光照时长对植物色泽和花青素的积累都有重要的影响.Lotkowska等[132 ] 发现苹果延长光照时间5 h后,上调了花青素合成转录因子MYB 112的表达,并诱导了苹果花青素的合成和积累.在茄子中分离出2个蓝光受体基因SmCRY 1、SmCRY 2和花青素调节阴性基因SmCOP 1和阳性基因SmHY 5,发现SmCRY 1、SmCRY 2和阳性基因SmHY 5在光照条件下的表达上调,而阴性基因SmCOP 1的表达下调[133 ] .另一方面,不同植物品种花青素合成积累对光照的响应也不同.利用RT-PCR技术分析FGM(非光敏型)和禾线(光敏型)两个光敏性不同品种茄子在套袋处理后果皮花青素合成酶基因和转录因子的表达情况,发现FGM与不套袋相比较,SmMYB 1的表达量显著增加,光形态建成因子SmCOP 1的表达量显著减少,而禾线与对照相比较,SmMYB 1 和SmCOP 1的表达情况则截然相反[134 ] .同时 Guan等[135 ] 以葡萄为试验材料,也得到了类似的结果,在光照条件下,Gamay葡萄的果皮和果肉中花青素含量显著多于Gamay Fre’aux. ...
Novel insight into the mechanism underlying light-controlled anthocyanin accumulation in eggplant (Solanum melongena L.)
1
2016
... 光照是非生物环境因子中影响花青素生物合成的关键因子,光照时长对植物色泽和花青素的积累都有重要的影响.Lotkowska等[132 ] 发现苹果延长光照时间5 h后,上调了花青素合成转录因子MYB 112的表达,并诱导了苹果花青素的合成和积累.在茄子中分离出2个蓝光受体基因SmCRY 1、SmCRY 2和花青素调节阴性基因SmCOP 1和阳性基因SmHY 5,发现SmCRY 1、SmCRY 2和阳性基因SmHY 5在光照条件下的表达上调,而阴性基因SmCOP 1的表达下调[133 ] .另一方面,不同植物品种花青素合成积累对光照的响应也不同.利用RT-PCR技术分析FGM(非光敏型)和禾线(光敏型)两个光敏性不同品种茄子在套袋处理后果皮花青素合成酶基因和转录因子的表达情况,发现FGM与不套袋相比较,SmMYB 1的表达量显著增加,光形态建成因子SmCOP 1的表达量显著减少,而禾线与对照相比较,SmMYB 1 和SmCOP 1的表达情况则截然相反[134 ] .同时 Guan等[135 ] 以葡萄为试验材料,也得到了类似的结果,在光照条件下,Gamay葡萄的果皮和果肉中花青素含量显著多于Gamay Fre’aux. ...
茄科蔬菜花青素苷分子调控研究进展
1
2022
... 光照是非生物环境因子中影响花青素生物合成的关键因子,光照时长对植物色泽和花青素的积累都有重要的影响.Lotkowska等[132 ] 发现苹果延长光照时间5 h后,上调了花青素合成转录因子MYB 112的表达,并诱导了苹果花青素的合成和积累.在茄子中分离出2个蓝光受体基因SmCRY 1、SmCRY 2和花青素调节阴性基因SmCOP 1和阳性基因SmHY 5,发现SmCRY 1、SmCRY 2和阳性基因SmHY 5在光照条件下的表达上调,而阴性基因SmCOP 1的表达下调[133 ] .另一方面,不同植物品种花青素合成积累对光照的响应也不同.利用RT-PCR技术分析FGM(非光敏型)和禾线(光敏型)两个光敏性不同品种茄子在套袋处理后果皮花青素合成酶基因和转录因子的表达情况,发现FGM与不套袋相比较,SmMYB 1的表达量显著增加,光形态建成因子SmCOP 1的表达量显著减少,而禾线与对照相比较,SmMYB 1 和SmCOP 1的表达情况则截然相反[134 ] .同时 Guan等[135 ] 以葡萄为试验材料,也得到了类似的结果,在光照条件下,Gamay葡萄的果皮和果肉中花青素含量显著多于Gamay Fre’aux. ...
茄科蔬菜花青素苷分子调控研究进展
1
2022
... 光照是非生物环境因子中影响花青素生物合成的关键因子,光照时长对植物色泽和花青素的积累都有重要的影响.Lotkowska等[132 ] 发现苹果延长光照时间5 h后,上调了花青素合成转录因子MYB 112的表达,并诱导了苹果花青素的合成和积累.在茄子中分离出2个蓝光受体基因SmCRY 1、SmCRY 2和花青素调节阴性基因SmCOP 1和阳性基因SmHY 5,发现SmCRY 1、SmCRY 2和阳性基因SmHY 5在光照条件下的表达上调,而阴性基因SmCOP 1的表达下调[133 ] .另一方面,不同植物品种花青素合成积累对光照的响应也不同.利用RT-PCR技术分析FGM(非光敏型)和禾线(光敏型)两个光敏性不同品种茄子在套袋处理后果皮花青素合成酶基因和转录因子的表达情况,发现FGM与不套袋相比较,SmMYB 1的表达量显著增加,光形态建成因子SmCOP 1的表达量显著减少,而禾线与对照相比较,SmMYB 1 和SmCOP 1的表达情况则截然相反[134 ] .同时 Guan等[135 ] 以葡萄为试验材料,也得到了类似的结果,在光照条件下,Gamay葡萄的果皮和果肉中花青素含量显著多于Gamay Fre’aux. ...
Anthocyanin biosynthesis is differentially regulated by light in the skin and flesh of white-fleshed and teinturier grape berries
1
2016
... 光照是非生物环境因子中影响花青素生物合成的关键因子,光照时长对植物色泽和花青素的积累都有重要的影响.Lotkowska等[132 ] 发现苹果延长光照时间5 h后,上调了花青素合成转录因子MYB 112的表达,并诱导了苹果花青素的合成和积累.在茄子中分离出2个蓝光受体基因SmCRY 1、SmCRY 2和花青素调节阴性基因SmCOP 1和阳性基因SmHY 5,发现SmCRY 1、SmCRY 2和阳性基因SmHY 5在光照条件下的表达上调,而阴性基因SmCOP 1的表达下调[133 ] .另一方面,不同植物品种花青素合成积累对光照的响应也不同.利用RT-PCR技术分析FGM(非光敏型)和禾线(光敏型)两个光敏性不同品种茄子在套袋处理后果皮花青素合成酶基因和转录因子的表达情况,发现FGM与不套袋相比较,SmMYB 1的表达量显著增加,光形态建成因子SmCOP 1的表达量显著减少,而禾线与对照相比较,SmMYB 1 和SmCOP 1的表达情况则截然相反[134 ] .同时 Guan等[135 ] 以葡萄为试验材料,也得到了类似的结果,在光照条件下,Gamay葡萄的果皮和果肉中花青素含量显著多于Gamay Fre’aux. ...
Tempranillo clones differ in the response of berry sugar and anthocyanin accumulation to elevated temperature
1
2018
... 对13个无性系葡萄品种分别进行24 ℃/14 ℃、28 ℃/18 ℃的处理,结果28 ℃下葡萄果皮花青素的积累量大于24 ℃下花青素的积累量[136 ] .对桃分别于16 ℃和12 ℃下贮藏后,发现在16 ℃下桃果皮存在花青素积累,而12 ℃或以下贮藏时花青素不积累,且16 ℃下桃果皮花青素生物合成酶基因PpPAL 1/2、PpC 4H 、Pp 4CL 、PpF 3H 、ppF 3'H、PpDFR 、PpANS 转录水平增加,促进了花青素的积累[137 ] .Liu等[138 ] 发现低温和低pH条件下有利于维持蓝莓总花色苷的最大稳定性.通过对生长在高温环境(30 ℃/15 ℃)下的草莓进行研究,发现其果皮和果肉中花青素浓度与对照(20 ℃/15 ℃)相比较均呈下降趋势,且转录因子FaMYB 10以及草莓花青素合成结构基因FaDFR 、FaANS 、FaUFGT 的表达均显著下调[139 ] .Ryu等[140 ] 通过下调和上调夜间温度来探讨夜间生长温度对苹果果皮色泽以及花青素积累的影响,结果发现降低夜温促进了苹果花青素生物合成基因MdCHS 、MdF 3H 、MdDFR 、MdANS 和MdUFGT 的表达,增加了花青素的积累,而上调夜温表现出相反的结果. ...
DNA demethylation is involved in the regulation of temperature-dependent anthocyanin accumulation in peach
1
2020
... 对13个无性系葡萄品种分别进行24 ℃/14 ℃、28 ℃/18 ℃的处理,结果28 ℃下葡萄果皮花青素的积累量大于24 ℃下花青素的积累量[136 ] .对桃分别于16 ℃和12 ℃下贮藏后,发现在16 ℃下桃果皮存在花青素积累,而12 ℃或以下贮藏时花青素不积累,且16 ℃下桃果皮花青素生物合成酶基因PpPAL 1/2、PpC 4H 、Pp 4CL 、PpF 3H 、ppF 3'H、PpDFR 、PpANS 转录水平增加,促进了花青素的积累[137 ] .Liu等[138 ] 发现低温和低pH条件下有利于维持蓝莓总花色苷的最大稳定性.通过对生长在高温环境(30 ℃/15 ℃)下的草莓进行研究,发现其果皮和果肉中花青素浓度与对照(20 ℃/15 ℃)相比较均呈下降趋势,且转录因子FaMYB 10以及草莓花青素合成结构基因FaDFR 、FaANS 、FaUFGT 的表达均显著下调[139 ] .Ryu等[140 ] 通过下调和上调夜间温度来探讨夜间生长温度对苹果果皮色泽以及花青素积累的影响,结果发现降低夜温促进了苹果花青素生物合成基因MdCHS 、MdF 3H 、MdDFR 、MdANS 和MdUFGT 的表达,增加了花青素的积累,而上调夜温表现出相反的结果. ...
Effect of temperature and pH on stability of anthocyanin obtained from blueberry
1
2018
... 对13个无性系葡萄品种分别进行24 ℃/14 ℃、28 ℃/18 ℃的处理,结果28 ℃下葡萄果皮花青素的积累量大于24 ℃下花青素的积累量[136 ] .对桃分别于16 ℃和12 ℃下贮藏后,发现在16 ℃下桃果皮存在花青素积累,而12 ℃或以下贮藏时花青素不积累,且16 ℃下桃果皮花青素生物合成酶基因PpPAL 1/2、PpC 4H 、Pp 4CL 、PpF 3H 、ppF 3'H、PpDFR 、PpANS 转录水平增加,促进了花青素的积累[137 ] .Liu等[138 ] 发现低温和低pH条件下有利于维持蓝莓总花色苷的最大稳定性.通过对生长在高温环境(30 ℃/15 ℃)下的草莓进行研究,发现其果皮和果肉中花青素浓度与对照(20 ℃/15 ℃)相比较均呈下降趋势,且转录因子FaMYB 10以及草莓花青素合成结构基因FaDFR 、FaANS 、FaUFGT 的表达均显著下调[139 ] .Ryu等[140 ] 通过下调和上调夜间温度来探讨夜间生长温度对苹果果皮色泽以及花青素积累的影响,结果发现降低夜温促进了苹果花青素生物合成基因MdCHS 、MdF 3H 、MdDFR 、MdANS 和MdUFGT 的表达,增加了花青素的积累,而上调夜温表现出相反的结果. ...
The effect of high air temperature on anthocyanin concentration and the expressions of its biosynthetic genes in strawberry ‘sachinoka’
1
2016
... 对13个无性系葡萄品种分别进行24 ℃/14 ℃、28 ℃/18 ℃的处理,结果28 ℃下葡萄果皮花青素的积累量大于24 ℃下花青素的积累量[136 ] .对桃分别于16 ℃和12 ℃下贮藏后,发现在16 ℃下桃果皮存在花青素积累,而12 ℃或以下贮藏时花青素不积累,且16 ℃下桃果皮花青素生物合成酶基因PpPAL 1/2、PpC 4H 、Pp 4CL 、PpF 3H 、ppF 3'H、PpDFR 、PpANS 转录水平增加,促进了花青素的积累[137 ] .Liu等[138 ] 发现低温和低pH条件下有利于维持蓝莓总花色苷的最大稳定性.通过对生长在高温环境(30 ℃/15 ℃)下的草莓进行研究,发现其果皮和果肉中花青素浓度与对照(20 ℃/15 ℃)相比较均呈下降趋势,且转录因子FaMYB 10以及草莓花青素合成结构基因FaDFR 、FaANS 、FaUFGT 的表达均显著下调[139 ] .Ryu等[140 ] 通过下调和上调夜间温度来探讨夜间生长温度对苹果果皮色泽以及花青素积累的影响,结果发现降低夜温促进了苹果花青素生物合成基因MdCHS 、MdF 3H 、MdDFR 、MdANS 和MdUFGT 的表达,增加了花青素的积累,而上调夜温表现出相反的结果. ...
Night temperatures affect fruit coloration and expressions of anthocyanin biosynthetic genes in ‘Hongro’ apple fruit skins
1
2017
... 对13个无性系葡萄品种分别进行24 ℃/14 ℃、28 ℃/18 ℃的处理,结果28 ℃下葡萄果皮花青素的积累量大于24 ℃下花青素的积累量[136 ] .对桃分别于16 ℃和12 ℃下贮藏后,发现在16 ℃下桃果皮存在花青素积累,而12 ℃或以下贮藏时花青素不积累,且16 ℃下桃果皮花青素生物合成酶基因PpPAL 1/2、PpC 4H 、Pp 4CL 、PpF 3H 、ppF 3'H、PpDFR 、PpANS 转录水平增加,促进了花青素的积累[137 ] .Liu等[138 ] 发现低温和低pH条件下有利于维持蓝莓总花色苷的最大稳定性.通过对生长在高温环境(30 ℃/15 ℃)下的草莓进行研究,发现其果皮和果肉中花青素浓度与对照(20 ℃/15 ℃)相比较均呈下降趋势,且转录因子FaMYB 10以及草莓花青素合成结构基因FaDFR 、FaANS 、FaUFGT 的表达均显著下调[139 ] .Ryu等[140 ] 通过下调和上调夜间温度来探讨夜间生长温度对苹果果皮色泽以及花青素积累的影响,结果发现降低夜温促进了苹果花青素生物合成基因MdCHS 、MdF 3H 、MdDFR 、MdANS 和MdUFGT 的表达,增加了花青素的积累,而上调夜温表现出相反的结果. ...
Anthocyanins as antidiabetic agents-In vitro and in silico approaches of preventive and therapeutic effects
1
2020
... 化合物的生物活性高度依赖于它们与环境的相互作用[141 ] .芸薹属蔬菜的生物活性与多种植物化学物质的活性有关,包括维生素C和E、类胡萝卜素、酚类化合物和硫代葡萄糖[142 ] .在红卷心菜中发现花青素是最丰富的酚类化合物[143 ] ,通过降低胆固醇含量有效保护心脏和肝脏. ...
Phytochemical composition and biological activities of differently pigmented cabbage (Brassica oleracea var. capitata) and cauliflower (Brassica oleracea var. botrytis) varieties
1
2019
... 化合物的生物活性高度依赖于它们与环境的相互作用[141 ] .芸薹属蔬菜的生物活性与多种植物化学物质的活性有关,包括维生素C和E、类胡萝卜素、酚类化合物和硫代葡萄糖[142 ] .在红卷心菜中发现花青素是最丰富的酚类化合物[143 ] ,通过降低胆固醇含量有效保护心脏和肝脏. ...
Antidiabetic effects and erythrocyte stabilization by red cabbage extract in streptozotocin-treated rats
1
2018
... 化合物的生物活性高度依赖于它们与环境的相互作用[141 ] .芸薹属蔬菜的生物活性与多种植物化学物质的活性有关,包括维生素C和E、类胡萝卜素、酚类化合物和硫代葡萄糖[142 ] .在红卷心菜中发现花青素是最丰富的酚类化合物[143 ] ,通过降低胆固醇含量有效保护心脏和肝脏. ...
Role of oxidative stress in development of complications in diabetes
1
1991
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
Free radicals and antioxidants in normal physiological functions and human disease
1
2007
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
Red cabbage anthocyanins: profile, isolation, identification, and antioxidant activity
1
2013
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
Antioxidant and pro-oxidant properties of acylated pelargonidin derivatives extracted from red radish (Raphanus sativus var. Niger , Brassicaceae )
1
2010
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
Characterization of purified red cabbage anthocyanins: improvement in HPLC separation and protective effect against H2 O2 -induced oxidative stress in HepG2 cells
1
2018
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
Antioxidant and antitumor effects and immunomodulatory activities of crude and purified polyphenol extract from blueberries
1
2016
... 人体代谢过程中产生多种自由基,而过多的自由基可导致脂质、蛋白质、DNA、RNA和糖的氧化[144 -145 ] .Teow等[21 ] 采用ORAC、DPPH和ABTS研究了19种甘薯白色、浅黄色、黄色、橙色和紫色果肉的自由基清除能力,结果表明紫色甘薯的抗氧化能力最强.这主要是因为花青素可以诱导各种氧化产物含量下降,防止氧化应激引起的胁迫损伤.Wiczkowski等[146 ] 探究了红甘蓝花青素的抗氧化活性,并报道了所有酰化的花青素苷比非酰化的花青素苷具有更强的抗氧化能力,此外,二酰基化的花青素比单酰基化的花青素具有更高的抗氧化能力.Wang等[147 ] 在对红萝卜花青素抗氧化性的研究中也得出类似结论.Fang等[148 ] 报道富红甘蓝花青素提取物可降低H2 O2 诱导的人体肝癌(HepG2)细胞氧化应激,改善细胞凋亡和细胞活力.Kou等[149 ] 在之前对蓝莓多酚提取物抗氧化性能分析的基础上,对粗提取物和纯化的花青素和多酚的抗氧化性能进行了探究,结果发现蓝莓花青素纯提取物具有较高的抗氧化活性. ...
紫甘薯花色苷对膀胱癌BIV87细胞增殖的影响及其机制探讨
1
2018
... 花青素和多酚类化合物对肿瘤的特殊抑制作用是近年来植物提取物治疗或辅助治疗癌症的研究热点.Zhao等[24 ] 的研究表明,小鼠体内植入S180肿瘤细胞后体重降低,然而紫甘薯花青素能够通过提高谷胱甘肽过氧化物酶和超氧化物歧化酶水平,增强抗氧化能力,丙二醛水平降低,从而抑制小鼠体内肿瘤生长;另一项关于紫甘薯的研究表明,花青素通过诱导细胞凋亡抑制膀胱癌BIU87细胞的增殖,且具有剂量依赖性[150 ] .此外,花青素对小鼠肿瘤生长的抑制率与花青素浓度成正比.槲皮素可抑制子宫肿瘤细胞的增殖,且其治疗效果优于其他活性物质.从葡萄中提取的花青素对乳腺癌MCF-7细胞的抗肿瘤作用优于纯化的花青素[151 ] ,这与Yang等[152 ] 的研究结果一致.另外,在一定程度上,活性物质对体外肿瘤增殖的抑制能力与抗氧化活性并不呈正相关[152 ] . ...
紫甘薯花色苷对膀胱癌BIV87细胞增殖的影响及其机制探讨
1
2018
... 花青素和多酚类化合物对肿瘤的特殊抑制作用是近年来植物提取物治疗或辅助治疗癌症的研究热点.Zhao等[24 ] 的研究表明,小鼠体内植入S180肿瘤细胞后体重降低,然而紫甘薯花青素能够通过提高谷胱甘肽过氧化物酶和超氧化物歧化酶水平,增强抗氧化能力,丙二醛水平降低,从而抑制小鼠体内肿瘤生长;另一项关于紫甘薯的研究表明,花青素通过诱导细胞凋亡抑制膀胱癌BIU87细胞的增殖,且具有剂量依赖性[150 ] .此外,花青素对小鼠肿瘤生长的抑制率与花青素浓度成正比.槲皮素可抑制子宫肿瘤细胞的增殖,且其治疗效果优于其他活性物质.从葡萄中提取的花青素对乳腺癌MCF-7细胞的抗肿瘤作用优于纯化的花青素[151 ] ,这与Yang等[152 ] 的研究结果一致.另外,在一定程度上,活性物质对体外肿瘤增殖的抑制能力与抗氧化活性并不呈正相关[152 ] . ...
Ultrasound-assisted enzymatic extraction of anthocyanins from grape skins: optimization, identification, and antitumor activity
1
2020
... 花青素和多酚类化合物对肿瘤的特殊抑制作用是近年来植物提取物治疗或辅助治疗癌症的研究热点.Zhao等[24 ] 的研究表明,小鼠体内植入S180肿瘤细胞后体重降低,然而紫甘薯花青素能够通过提高谷胱甘肽过氧化物酶和超氧化物歧化酶水平,增强抗氧化能力,丙二醛水平降低,从而抑制小鼠体内肿瘤生长;另一项关于紫甘薯的研究表明,花青素通过诱导细胞凋亡抑制膀胱癌BIU87细胞的增殖,且具有剂量依赖性[150 ] .此外,花青素对小鼠肿瘤生长的抑制率与花青素浓度成正比.槲皮素可抑制子宫肿瘤细胞的增殖,且其治疗效果优于其他活性物质.从葡萄中提取的花青素对乳腺癌MCF-7细胞的抗肿瘤作用优于纯化的花青素[151 ] ,这与Yang等[152 ] 的研究结果一致.另外,在一定程度上,活性物质对体外肿瘤增殖的抑制能力与抗氧化活性并不呈正相关[152 ] . ...
Investigation on the biological activity of anthocyanins and polyphenols in blueberry
2
2021
... 花青素和多酚类化合物对肿瘤的特殊抑制作用是近年来植物提取物治疗或辅助治疗癌症的研究热点.Zhao等[24 ] 的研究表明,小鼠体内植入S180肿瘤细胞后体重降低,然而紫甘薯花青素能够通过提高谷胱甘肽过氧化物酶和超氧化物歧化酶水平,增强抗氧化能力,丙二醛水平降低,从而抑制小鼠体内肿瘤生长;另一项关于紫甘薯的研究表明,花青素通过诱导细胞凋亡抑制膀胱癌BIU87细胞的增殖,且具有剂量依赖性[150 ] .此外,花青素对小鼠肿瘤生长的抑制率与花青素浓度成正比.槲皮素可抑制子宫肿瘤细胞的增殖,且其治疗效果优于其他活性物质.从葡萄中提取的花青素对乳腺癌MCF-7细胞的抗肿瘤作用优于纯化的花青素[151 ] ,这与Yang等[152 ] 的研究结果一致.另外,在一定程度上,活性物质对体外肿瘤增殖的抑制能力与抗氧化活性并不呈正相关[152 ] . ...
... [152 ]. ...
Polyphenol-rich bilberry ameliorates total cholesterol and LDL-cholesterol when implemented in the diet of Zucker diabetic fatty rats
1
2013
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Red cabbage (Brassica oleracea L.) mediates redox-sensitive amelioration of dyslipidemia and hepatic injury induced by exogenous cholesterol administration
1
2014
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Anthocyanin-rich Riceberry bran extract attenuates gentamicin-induced hepatotoxicity by reducing oxidative stress, inflammation and apoptosis in rats
1
2017
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Effects of anthocyanins on psychological stress-induced oxidative stress and neurotransmitter status
1
2008
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Anthocyanins improve hippocampus-dependent memory function and prevent neurodegeneration via JNK/akt/GSK3β signaling in LPS-treated adult mice
1
2019
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Anthocyanins protect against LPS-induced oxidative stress-mediated neuroinflammation and neurodegeneration in the adult mouse cortex
1
2016
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Anthocyanins reversed D-galactose-induced oxidative stress and neuroinflammation mediated cognitive impairment in adult rats
1
2017
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Anthocyanins isolated from blueberry ameliorates CCl4 induced liver fibrosis by modulation of oxidative stress, inflammation and stellate cell activation in mice
1
2018
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
Anti-hyperglycemic effect of diacylated anthocyanin derived from Ipomoea batatas cultivar Ayamurasaki can be achieved through the alpha-glucosidase inhibitory action
1
2002
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
The modulatory effect of anthocyanins from purple sweet potato on human intestinal microbiota in vitro
1
2016
... 来自医学研究的试验数据表明,含有花青素的提取物或纯花青素对保护心脏具有巨大的潜力.Brader等[153 ] 研究了富含花青素的蔓越莓和黑加仑浆果的饮食(每天5 g含有172 mg花青素的浆果粉)对糖尿病脂肪大鼠的脂质特征和其他生物指标的影响.实验进行56 d后的结果显示,胆固醇水平降低,有效保护动物体肝脏.Al-Dosari等[154 ] 发现,高剂量的红白菜提取物可以改善大鼠因胆固醇积累导致的肝组织的坏死、炎症和纤维化.Arjinajarn等[155 ] 研究发现富含矢车菊素-3-O-葡萄糖苷和芍药苷-3-O-葡萄糖苷(分别为13.24和5.33 mg·g-1 粗提物)的米糠提取物可有效预防庆大霉素引起的肝脏中毒.此外,口服富含花青素的越橘浆果提取物(100 mg·kg-1 ,连续7 d)可抑制心理应激诱导的小鼠大脑氧化应激和多巴胺异常[156 ] .从黑大豆中提取的花青素也被证明可以逆转成年小鼠中d-半乳糖或脂聚糖诱导的氧化应激、神经炎症和神经变性[157 ⇓ -159 ] .另外,在研究过程中还发现花青素处理后,恢复了线粒体电子传递链复合物Ⅰ和Ⅱ的活性[160 ] .Matsui等[161 ] 先给予8周龄雄性小鼠2 g·kg-1 BW的麦芽糖,然后灌胃注入100 mg·kg-1 BW的二酰化紫甘薯花青素,结果显示30 min后小鼠血糖浓度明显下降16.5%,二酰化花青素通过抑制α-葡萄糖苷酶活性降低血糖水平.Zhang等[162 ] 通过观察紫甘薯花青素处理后不同时间点细菌数量和短链脂肪酸浓度的变化,分析其对肠道菌群的影响.结果发现,花青素能明显增加双歧杆菌和乳酸杆菌/肠球菌的种群数量和浓度.因此,摄入富含花青素的紫甘薯有利于肠道微生态和宿主健康. ...
A CONSORT-compliant, randomized, double-blind, placebo-controlled pilot trial of purified anthocyanin in patients with nonalcoholic fatty liver disease
1
2015
... 在人体水平上,Zhang等[163 ] 报道,从黑加仑中提取花青素(每天320 mg)用于临床治疗84 d后,可改善非酒精性脂肪肝患者的肝损伤.在另一项研究中,测试了含有高多酚和花青素含量的果汁(40%红葡萄汁、20%黑莓汁、15%酸樱桃汁、15%黑加仑汁和10%接骨木果汁)对血液透析患者的预防作用,结果表明,DNA氧化损伤、蛋白质和脂质过氧化水平显著降低,还原型谷胱甘肽水平升高[164 ] .血液透析患者经常饮用浓缩红葡萄汁可通过降低中性粒细胞NADPH氧化酶活性以及炎症标志物水平降低进而缓解病症[165 ] . ...
Anthocyanin/polyphenolic-rich fruit juice reduces oxidative cell damage in an intervention study with patients on hemodialysis
1
2008
... 在人体水平上,Zhang等[163 ] 报道,从黑加仑中提取花青素(每天320 mg)用于临床治疗84 d后,可改善非酒精性脂肪肝患者的肝损伤.在另一项研究中,测试了含有高多酚和花青素含量的果汁(40%红葡萄汁、20%黑莓汁、15%酸樱桃汁、15%黑加仑汁和10%接骨木果汁)对血液透析患者的预防作用,结果表明,DNA氧化损伤、蛋白质和脂质过氧化水平显著降低,还原型谷胱甘肽水平升高[164 ] .血液透析患者经常饮用浓缩红葡萄汁可通过降低中性粒细胞NADPH氧化酶活性以及炎症标志物水平降低进而缓解病症[165 ] . ...
Comparative effects of dietary supplementation with red grape juice and vitamin E on production of superoxide by circulating neutrophil NADPH oxidase in hemodialysis patients
1
2008
... 在人体水平上,Zhang等[163 ] 报道,从黑加仑中提取花青素(每天320 mg)用于临床治疗84 d后,可改善非酒精性脂肪肝患者的肝损伤.在另一项研究中,测试了含有高多酚和花青素含量的果汁(40%红葡萄汁、20%黑莓汁、15%酸樱桃汁、15%黑加仑汁和10%接骨木果汁)对血液透析患者的预防作用,结果表明,DNA氧化损伤、蛋白质和脂质过氧化水平显著降低,还原型谷胱甘肽水平升高[164 ] .血液透析患者经常饮用浓缩红葡萄汁可通过降低中性粒细胞NADPH氧化酶活性以及炎症标志物水平降低进而缓解病症[165 ] . ...
A simple and rapid analytical method of grape anthocyanin pigments by cellulose thin-layer chromatography and densitometry
2
1979
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Separation of anthocyanin chalcones by high-performance liquid chromatography
1
1981
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Application of RP-HPLC and spectrophotometry in standardization of bilberry anthocyanin extract
2
1992
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Effect of enzyme-aided pressing on anthocyanin yield and profiles in bilberry and blackcurrant juices
1
2005
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Isolation and structure charactrisation of anthocyanin pigments in black carrot (Daucus carota L.)
2
2006
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Simultaneous separation by reversed-phase high-performance liquid chromatography and mass spectral identification of anthocyanins and flavonols in Shiraz grape skin
1
2008
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Activity and contents of polyphenolic antioxidants in the whole fruit, flesh and peel of three apple cultivars
2
2009
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Metabolite fingerprinting of bokbunja (Rubus coreanus miquel) by UPLC-qTOF-MS
2
2011
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Liquid chromatography/mass spectrometry based fingerprinting analysis and mass profiling of Euterpe oleracea (açaí) dietary supplement raw materials
1
2012
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Herbal mouthwash: a review
2
2022
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Protective effect of blueberry anthocyanins in a CCl4 -induced injury model in human embryonic liver cells
1
2014
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
A multiplex approach for the UPLC-PDA-MS/MS data: analysis of licorice
1
2014
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
定量核磁共振法测定山楂提取物中的原花青素
2
2015
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
定量核磁共振法测定山楂提取物中的原花青素
2
2015
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Identification and quantitation of anthocyanins in purple-fleshed sweet potatoes cultivated in China by UPLC-PDA and UPLC-QTOF-MS/MS
2
2016
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Ohmic heating-assisted extraction of anthocyanins from black rice bran to prepare a natural food colourant
1
2015
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
ABDEL SHAKOUR Z T. Metabolomics driven analysis of 11 Portulaca leaf taxa as analysed via UPLC-ESI-MS/MS and chemometrics
2
2019
... Analytical method of anthocyanin
Table 3 物种 测定样品/部位 方法 参考文献 葡萄Vitis vinifera L. 果肉Pulp 纤维素薄层色谱-密度分析法Thin-layer chromatography [166 ] 葡萄Vitis vinifera L. 果皮Pericarp 高效液相色谱法High-performance liquid chromatography [167 ] 越橘Vaccinium dunalianum 叶片Leaf 紫外分光光度计法The UV spectrophotometry [168 ] 蓝莓Vaccinium spp. 果实Fruit 酶辅助提取Enzyme-assisted extraction [169 ] 黑胡萝卜Daucus carota L. 根Root pH示差法The pH-differential spectrophotometry [170 ] 葡萄Vitis vinifera L. 果皮Pericarp 反向高效液相色谱法 [171 ] Reversed phase high-performance liquid chromatography 苹果Malus ×domestica 果实Fruit 对-二甲基氨基肉桂醛法The P-dimethylamino-cinnamaldehyde method [172 ] 覆盆子Rubus idaeus L. 果实Fruit 超高效液相色谱-光电二极管阵列检测器-四极杆飞行时间质谱法 [173 ] UPLC-PDA-Q/TOF-MS 巴西莓 冻干巴西莓粉 液相色谱-质谱联用测定法LS-MS [174 ] Euterpe oleracea Mart.Freeze-dried acaie powder 蔓越莓Oxycoccos 果肉Pulp 香草醛法Vanillin method [175 ] 蓝莓Vaccinium spp. 果肉Pulp 电化学发光法Electrochemiluminescence [176 ] 甘草 根Root 超高效液相色谱-二极管阵列检测器串联质谱法MS/MS- UPLC-PDA [177 ] Glycyrrhiza uralensis Fisch. 山楂 果肉Pulp 定量核磁共振法Quantitative NMR [178 ] Crataegus pinnatifida Bge. 马铃薯Solanum tuberosum 块茎Tuber 超高效液相色谱-二极管阵列检测器法UPLC-PDA [179 ] 黑稻Oryza sativa L. 种子Seeding 欧姆加热辅助萃取Ohmic heating assisted extraction [180 ] 马齿苋Portulaca oleracea L. 叶片Leaf 超高效液相色谱-电喷雾串联三重四级杆质谱法UPLC-ESI-MS/MS [181 ]
紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
pH示差法测定蓝莓酒中花色苷条件的优化
1
2011
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
pH示差法测定蓝莓酒中花色苷条件的优化
1
2011
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
Application of quantitative1 H-NMR method to determination of gentiopicroside in Gentianae radix and Gentianae scabrae radix
1
2014
... 紫外分光光度计法是测定花青素含量简便且成本较低的办法[168 ] .目前,高效液相色谱法是分离、提取.在高效液相色谱法测定植物花青素含量的基础上,超高效液相色谱-二极管阵列检测器串联质谱定性分析法(MS/MS-UPLC-PDA)、超高效液相-四级杆飞行时间质谱联用法(UPLC-PDA-Q/TOF-MS)以及超高液相色谱-电喷雾二级质谱联用(UPLC-ESI-MS/MS)等方法被证明具有更精确、重复性更好的特点[173 ,179 ,181 ] .有研究发现,植物花青素糖苷结构会随着pH值的变化而变化,而其他会造成测定干扰的物质的特征光谱并不会在环境条件的影响下改变,因此,在样品中含有干扰物时,要测定花青素总量选用pH值示差法是比较合适的测定方法[170 ,182 ] .此外,也有利用花青素化学结构而添加对-二甲基氨基肉桂醛或香草醛等使其官能团发生化学反应,以此形成稳定的花青素缩合产物,然后用成本较低、简便的分光光度计法测定花青素物质的量浓度[172 ,175 ] .为了快速检测大量样品中的特征物质,用定量核磁共振法测定花青素种类和含量,具有分析速度快,精确度高的特点[178 ,183 ] .另外,纤维素薄层色谱-密度分析法也被常用于分离和定性分析花青素的测定中,对其测定过程中杂质以及干扰物质的鉴定具有重要的作用[166 ,184 ] . ...
薄层及凝胶色谱法测定树莓籽原花青素的平均聚合度
0
2013
薄层及凝胶色谱法测定树莓籽原花青素的平均聚合度
0
2013


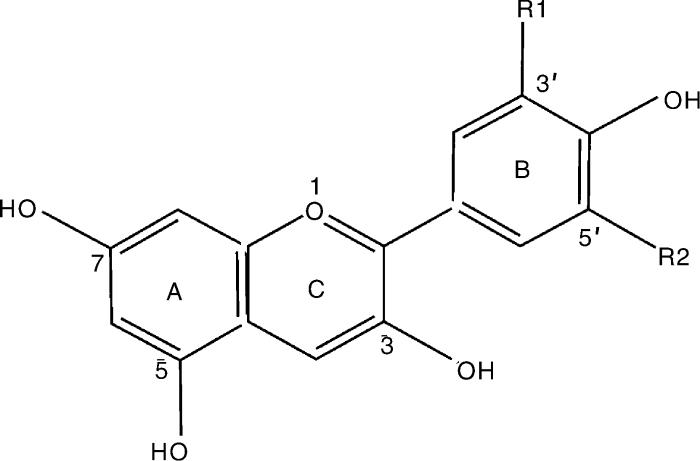
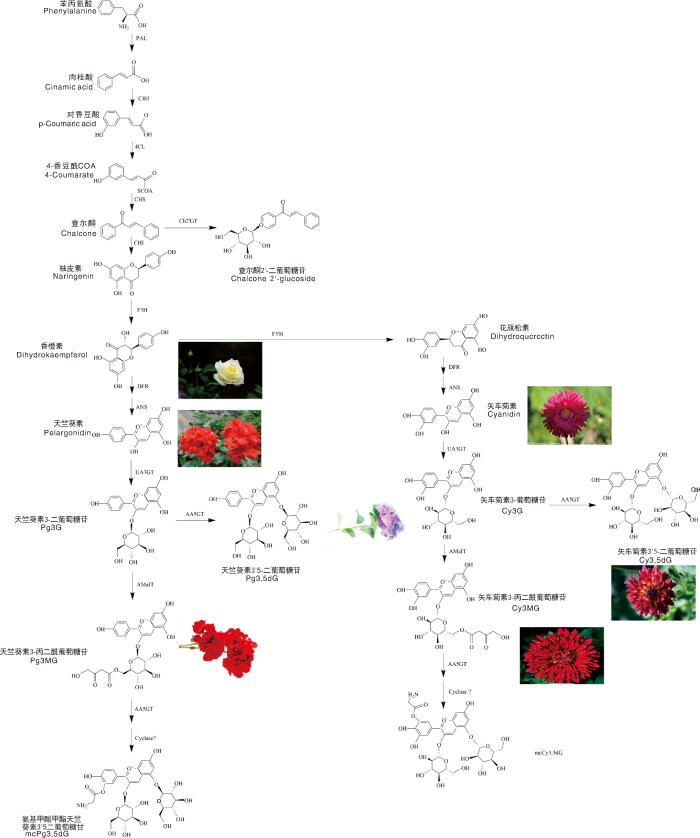


{kind=link}
{kind=link}
{kind=link}
{kind=link}